Главная Обратная связь
Дисциплины:
Архитектура (936)
Биология (6393)
География (744)
История (25)
Компьютеры (1497)
Кулинария (2184)
Культура (3938)
Литература (5778)
Математика (5918)
Медицина (9278)
Механика (2776)
Образование (13883)
Политика (26404)
Правоведение (321)
Психология (56518)
Религия (1833)
Социология (23400)
Спорт (2350)
Строительство (17942)
Технология (5741)
Транспорт (14634)
Физика (1043)
Философия (440)
Финансы (17336)
Химия (4931)
Экология (6055)
Экономика (9200)
Электроника (7621)

Зрительное опознание жертвы и хищника
|
|
Большинство хищников встречается с множеством разных видов, которые служат им добычей и которых необходимо отличать от других организмов. При этом чаще всего используются три признака - величина, движение и форма.
Когда хищники должны выбирать между особями-жертвами, отличающимися только размерами тела, они обычно хватают самую крупную. Эта стратегия наиболее эффективна в отношении затрат энергии. Однако по мере увеличения размера обычно наступает предел, за которым стимул больше не воспринимается как добыча. Например, когда обыкновенной жабе (Bufo bufo) предъявляют как «добычу» объекты разной величины, она реагирует положительно на экземпляры, размеры которых находятся в определенных пределах, но активно избегает более крупные стимулы (рис. 13.6).
Как жаба судит о величине объекта? Проще всего делать это по величине изображения на сетчатке, измеряемого градусами угла зрения. Для объекта постоянной величины угол зрения меняется с расстоянием объекта от глаза. Близкие объекты выглядят крупнее, чем удаленные. Чтобы выбрать добычу определенной величины, жабе нужно оценить абсолютную величину видимого объекта, учитывая и его размеры на сетчатке, и расстояние до него. Жабы-повитухи (Alytes obstetricans] в процессе развития постепен-

| Рис. 13.6. Жабы реагируют на маленькие движущиеся квадраты, ориентируясь в их направлении, но при определенных размерах квадрата эта реакция исчезает и большие квадраты вызывают реакцию избегания. (По Ewert, 1980.) |
но научаются выбирать добычу постоянного размера (Ewert, Burghagen, 1979). Сразу после метаморфоза они предпочитают искусственную приманку некоторого углового размера почти независимо от расстояния. Через шесть месяцев они ориентируются на объекты определенного абсолютного размера независимо от расстояния (рис. 13.7), т.е. за это время они как-то научаются включать суждения о расстоянии в свои оценки размеров. В некоторых случаях наблюдается склонность к более крупной добыче, но не из-за активного предпочтения, а потому, что она лучше различима. Так, радужные форели (Salmo gairdneri) чаще ловят крупных ракообразных, чем мелких, потому что крупные видны на большем расстоянии (Ware, 1972).
Для того чтобы узнать добычу, некоторым видам, например лягушкам и жабам, нужно, чтобы она двигалась. Обыкновенная каракатица (Sepia officinalis) в норме нападает только на движущихся креветок. Но если у каракатицы отнять только что пойманную и парализованную ею креветку, то это головоногое немедленно снова нападет на нее даже на неподвижную (Messenger, 1968). Некоторые хищники предпочитают добычу, которая движется беспорядочно. Так, личинки стрекоз предпочитают животных, совершающих зигзагообразные движения (Etienne, 1969), а солнечная рыба (Lepomis gibbosus) скорее нападет на искусственную рыбку, если та извивается, чем на плавно движущийся объект (Gandolfi et al., 1968). Иногда имеет значение соотношение формы и движения. Так, когда жабам предъявляют темную движущуюся полосу на белом фоне, они сразу же нападают на нее, когда полоса перемещается вдоль своей оси подобно червяку (рис. 13.8).
Узнавание добычи по форме - настолько сложное дело, что тут трудно сформулировать какие-нибудь общие правила. В ряде поведенческих работ Робинсон (Robinson, 1970) давал в неволе насекомых тамаринам (Saguinus geoffroyi) - восновном насекомоядным обезьянам Нового Света. В некоторых опытах им предъявляли богомолов - нормального, без головы, с двумя головами и т.д. Результаты показывают, что для узнавания добычи этим обезьянам важно было опознать голову. В опытах с палочниками в качестве приманки тамарины склонны были пренебрегать насекомыми без ног, но быстро хватали экземпляры с хорошо заметными ногами или маленькие палочки с прикрепленными к ним ножками насекомых. Таким образом, для опознания добычи тамаринам важны головы и конечности насекомых. Чтобы спастись от хищника, насекомые-жертвы выработали разнообразные защитные приспособления, скрывающие конечности и искажающие очертания тела. В опытах с абстрактными

|
| Рис. 13.7. Молодые жабы (вверху) оценивают размеры по углу зрения, а взрослые жабы (внизу) могут судить о размерах независимо от угла зрения. (По Ewert, 1980.) |
двумерными фигурами Робинсон обнаружил, что двусторонняя симметрия, вероятно, служит одним из признаков добычи, на который обычно ориентированы хищники.
Те же принципы применимы и к узнаванию жертвами хищников. Так, например, силуэт ястреба, движимый над утятами или гусятами, вызывает реакцию страха, когда движется только в определенном направлении (рис. 13.9). Это объясняется тем, что короткая шея и длинный хвост характерны для ястреба, а длинная шея и короткий хвост - признаки летящего гуся (Tinbergen, 1951). Жабы избегают фигур, похожих на змею с поднятой головой (рис. 13.10). Пиявка, движущаяся толчками, воспринимается как добыча, если ее передняя присоска находится на почве, но если эта присоска поднята в воздух, жаба принимает пиявку за врага (Ewert, 1980).
Как мы видим, поведенческие исследования (см. обзор Ewert, 1980) показывают, что жабы хватают мелкие продолговатые объекты, расположенные горизонтально, но не реагируют на такие же объекты в вертикальном положении (рис. 13.8). Жаба обычно питается насекомыми, личинками, червяками и т. п. Ее хищническое поведение состоит из ориентации головы и туловища, зрительной фиксации добычи, захватывания ее в результате вытягивания шеи и языка, глотания и вытирания морды передними лапами (рис. 13.11). Чтобы вызвать хищническое поведение, нужен мелкий движущийся объект. На крупные движущиеся объекты жаба отвечает оборонительным поведением. Хотя для изучения сенсорных процессов могут быть применены разные чисто поведенческие методы, физиологическое исследование тоже может давать ценные сведения о деятельности органов чувств и о типе информации, которую они посылают в мозг. Но для того чтобы установить, как мозг использует такую информацию, тре-

| Рис. 13.8. Жабы реагируют положительно на прямоугольники, движущиеся вперед узкой стороной, и отрицательно на движущиеся вперед широкой стороной. (По Ewert, 1980.) |

| Рис. 13.9. Этот силуэт похож на ястреба, если его двигать в одном направлении, и на гуся, если его двигать в противоположном направлении. |

|
| Рис. 13.10. Реакции жабы на простые модели: А - змея; Б - абстрактный рисунок; В - пиявка с поднятой головой; Г - пиявка с опущенной головой. Первые три вызывают реакцию избегания, последняя исследуется. (По Ewert, 1980.) |
буется сочетание поведенческого и физиологического подходов. Такой подход был применен Эвертом и его коллегами в обширных исследованиях по опознанию добычи и врагов жабами.
Физиологические работы показывают, что некоторое опознание добычи происходит на уровне сетчатки. Леттвин и др. (Lettvin et al., 1959) отводили электрическую активность от зрительного нерва лягушки, когда объекты двигались в поле зрения. Они обнаружили четыре типа ответов, которые, по-видимому, соответствуют четырем типам ганглиозных клеток в сетчатке. Было установлено, что эти клетки являются детекторами: 1) неподвижной границы, 2) темного выпуклого движущегося объекта, 3) изменения контраста или движения и 4) затемнения. Жаба, очевидно, обладает тремя типами ганглиозных клеток (соответствующих типам 2, 3 и 4 у лягушки), аксоны которых в составе зрительного нерва идут в зрительную покрышку головного мозга (Ewert, Hock, 1972). Информация, поступающая в мозг, включает угловой размер и скорость движения объекта, степень контраста с фоном и общий уровень освещенности. Однако, чтобы распознать добычу, жабе этой информации недостаточно.
Методом дегенерации можно проследить путь волокон зрительного нерва в разные отделы головного мозга, включая зрительную покрышку и претектальную область таламуса (рис. 13.12). Сетчатка одного глаза проецируется топографически на поверхностные слои противоположной зрительной покрышки. Эта проекция имеет вид карты, на которой каждая область поля зрения соответствует определенной области зрительной покрышки. Электрическая стимуляция покрышки у свободно движущейся жабы приводит к ориентировочной реакции по направлению к соответствующей части поля зрения, т. е. вызывает такое поведение, как будто соответствующая часть зрительной покрышки стимулируется видом добычи. Электрическая стимуляция проекции сетчатки на таламической претектальной области вызывает у жабы реакцию избегания.

|
| Рис. 13.11. Обнаружение и схватывание добычи жабой. Сверху вниз: добыча попадает в боковое поле зрения животного; жаба поворачивается к добыче, подводя ближе к ней центр поля зрения, а в результате используя бинокулярное поле; затем жаба схватывает добычу языком. (По Ewert, 1980.) |

| Рис. 13. 12. Соотношение между частями головного мозга жабы, участвующими в регуляции избегания и в ориентировочной реакции на зрительно обнаруживаемые обьекты. (По Ewert, 1980.) |
Хирургическое разрушение части головного мозга (претектальной области таламуса) приводит к тому, что животные начинают хватать любой движущийся предмет. Разрушение зрительной покрышки уничтожает всякую реакцию на движущиеся стимулы, в том числе и поведение избегания. Эти данные побудили Эверта (Ewert, 1980) постулировать, что проекция сетчатки на претектальную область таламуса вызывает поведение избегания, но что для этого также требуется некоторый приток возбуждения от зрительной покрышки. Проекция сетчатки на зрительную покрышку служит основой реакции схватывания всех движущихся объектов, но реакция на крупные или похожие на врага объекты тормозится претектальной областью таламуса и схватываются только мелкие объекты.
Гипотеза Эверта подтверждается физиологическими работами по изучению электрической активности нейронов в зрительной покрышке и претектальной области таламуса в ответ на стимуляцию сетчатки и других связанных с этой функцией частей головного мозга (Ewert, 1980). Это исследование представляет большой интерес не только как демонстрация соответствующего участия сетчатки и мозга в фильтрации стимула, но так же как прекрасный пример того, чего можно достичь разумным сочетанием поведенческих и физиологических методов.
ДЛЯ ЗАПОМИНАНИЯ
1. Животные, обитающие при слабом освещении, часто обладают специальными зрительными приспособлениями - глазами цилиндрической формы, тапетумом и зрительными пигментами, - предназначенными для повышения чувствительности в этих условиях.
2. У животных, которые живут или активны в темноте, не позволяющей видеть окружающие предметы, глаза могут дегенерировать и зрение заменяют другие сенсорные системы. К ним относятся эхолокационная система летучих мышей, система слуховой локализации у сов и электрическая чувствительность у некоторых рыб.
3. Зрительное опознание добычи и хищников часто включает в себя обнаружение ключевых раздражителей, что позволяет животному быстро принимать решение и быстро реагировать.
Рекомендуемая литература
Gould J. L. (1980) The case for magnetic-field sensitivity in birds and
bees (such as it is). American Scientist, 68,256-267.
Lythgoe J.N. (1979) The Ecology of Vision, Clarendon, Oxford.
II.2. Животное и среда
В этом разделе будут рассмотрены механизмы, используемые животными для регуляции их отношений со средой. Гл. 14 начинается с анализа координации и ориентации тела и переходит от вопросов простой ориентации к навигации. Гл. 15 рассматривает внутреннюю среду животного. Понятие гомеостаза разбирается в связи с терморегуляцией, питанием и питьем. В гл. 16 показано, как животное может приспособить свою физиологию и поведение к изменениям среды. Достаточно подробно обсуждается роль биологических часов с точки зрения физиологии размножения, зимней спячки, миграции и суточной активности.
Клод Бернар (1813-1878)

|
| (ВВС Ниlton Portrate Library). |
Клод Бернар изучал медицину с 1834 по 1843 г., а затем занялся исследованиями в Коллеж де Франс как ученик Мажанди. Его главные открытия сделаны между 1840 и 1859 гг., после чего он мало занимался экспериментальной работой, а посвятил себя разработке своих теорий и писательской деятельности.
Основная работа Бернара "Introduction a 1'Etude de la medicine experimentale" («Введение в изучение экспериментальной медицины») издана в 1865 г. До этого он выпускал различные "Lecons" («Лекции»), но они обычно представляли собой записи, сделанные его учениками на лекциях и опубликованные под его наблюдением. Бернару принадлежат многие другие работы, вершиной которых является труд "Lecons sur les Phenomienes de la Vie, Communs aux Animaux et aux Vegetaux" («Лекции о жизненных явлениях, обших для животных и растений»), вышедший в свет вскоре после его смерти в 1878 г.
До трудов Бернара тело считали совокупностью органов, каждый из которых выполняет независимые от других функции. Клод Бернар показал, что разные виды физиологической активности взаимосвязаны и организм следует рассматривать как единую сложную и совершенную машину.
С 1848 г., работая в Коллеж де Франс, Клод Бернар проверял действие на животных разной пищи. Он обнаружил, что их кровь всегда содержит сахар независимо от того, когда он съеден. Даже у собак, получавших только мясо, в крови был сахар, который, как показал Клод Бернар, попадал в кровь из печени. Он установил, что печень накапливает некоторые углеводы, получаемые при переваривании пищи, и выделяет сахар в кровь, когда его нет в рационе. Клод Бернар провел также важные исследования пищеварения и получил данные о регуляции кровоснабжения разных частей организма.
Клода Бернара считают основателем современной экспериментальной физиологии. Одно из его часто цитируемых изречений гласит: «Зачем думать, когда можно экспериментировать? Исчерпайте эксперимент и тогда думайте». Его экспериментальная работа всегда была направлена на проверку определенной гипотезы. Комментируя «блестящую работу Лапласа» и указав, что биологи мало используют статистику, он заметил: «Разумеется, я думаю, что через каких-нибудь сто лет все будут не только употреблять статистику, но и злоупотреблять ею и будут полагаться на этот метод для спасения работы, предпринятой без связи с какой-либо рабочей гипотезой».
Тем не менее основная заслуга Клода Бернара состоит в создании новых теоретических концепций. Его самое известное изречение «Постоянство внутренней среды - условие свободной и независимой жизни» впервые появилось в 1859 г. в одной из «Лекций» и затем было развито в его посмертном труде 1878 г.Он понял, что кровь служит внутренней средой для всех тканей организма и что на состояние крови влияют отчасти внутренние процессы и отчасти изменения внешней среды. Животные, способные поддерживать постоянство внутренней среды, обладают гораздо большей экологической свободой, чем животные, зависящие от колебаний внешних условий. Идеей о регуляции внутренней среды была проникнута вся научная деятельность Клода Бернара, хотя ученые стали подробно обсуждать эту концепцию только в следующем столетии. Она оказала глубокое влияние на современные представления о физиологии и поведении животных.
Координация и ориентация
В этой главе будет рассказано о пространственных отношениях животного с его внешней средой. В ответ на изменения среды животные должны координировать и ориентировать свои движения по отношению к внешним стимулам. Сначала мы рассмотрим координацию, а затем ориентацию и закончим обсуждением навигации — самой сложной формы пространственной ориентации.
Координация
Как показано в гл. И, эффективная координация движения и локомоции зависят от информации, получаемой ЦНС о положении, напряжении и других состояниях мышц; эта информация поступает от внутренних органов чувств - мышечных веретен, сухожильных органов и суставных рецепторов. Кроме передачи информации об относительном положении конечностей и других органов движения эти органы чувств позволяют животным давать рефлекторные ответы на изменения, вызываемые внешними агентами или самим движением животного.
Рефлекторное поведение - самая простая форма реакции на стимулы. В норме рефлексы бывают автоматическими, непроизвольными и стереотипными. Внезапное изменение напряжения мышцы может вызвать автоматическую смену позы, а резкое изменение уровня освещенности - рефлекторное отстранение. Рефлексы могут быть относительно локальными, т.е. затрагивать одну конечность или какую-либо другую часть тела, как показано на рис. 14.1, но иногда в рефлекторную реакцию вовлекается все животное. Например, реакция вздрагивания у человека требует рефлекторной координации многих мышц. Точно так же рефлекс отстранения у беспозвоночных - полихет и различных моллюсков - охватывает все тело. Как было показано в гл. 11, такие рефлексы запускаются гигантскими нервными волокнами, которые очень быстро передают сигналы всем участвующим в ответе мышцам, так что они сокращаются сразу и одновременно.
Рефлексы у позвоночных в норме автоматические, однако нередко они подвержены различным воздействиям через центральные синапсы (рис. 14.1). Так, с рефлексами позы часто взаимодействуют другие, несовместимые с ними рефлекторные механизмы. Если в двух рефлексах используются одни и те же мышцы, то эти рефлексы несовместимы, т.е. не могут протекать одновременно. Такие пары рефлексов несовместимы также неврологически в том смысле, что стимуляция одного рефлекса тормозит другой. Торможение обычно бывает реципрокным и составляет самую элементарную форму координации, когда одна активность полностью подавляет другую или подавляется ею. Этот тип реципрокного торможения характерен для ходьбы и других видов локомоции.
Вообще мышечная координация определяется двумя главными процессами - центральной и периферической регуляциями. При центральной регуляции головной мозг дает точные команды, которые выполняются соответствующими мышцами. К такому типу, по-видимому, относится координация глотательных движений у млекопитающих (Doty, 1968). Центральная регуляция важна для координации многих выученных движений, требующих

| Рис. 14.1. Схема простого коленного рефлекса. Удар под коленной чашкой стимулирует рецепторы в сухожилии, которые посылают сигналы в спинной мозг. Сигналы передаются двигательным нервам, управляющим мышцами-разгибателями, которые рефлекторно сокращаются, подбрасывая ногу. |
быстрой мышечной активности. Так, например, каракатица Sepia ловит мелких ракообразных двумя вытягивающимися щупальцами (рис. 14.2). Регуляция нападения делится на две фазы. Первая представляет собой направляемую зрением систему, в которой добыча приводится бинокулярно в фокус, и движения каракатицы следуют за движениями жертвы таким образом, что ошибка «прицеливания» сводится к нулю. Когда фаза слежения завершена, щупальцы внезапно выбрасываются и схватывают жертву. Завершающая фаза такая быстрая (около 30 мс), что времени на зрительный контроль движения не остается. Это можно показать, выключив свет во время выбрасывания щупалец: все равно жертва будет поймана. Но если в это время добыча переместится, то каракатица ее не поймает; значит, это головоногое не способно корректировать свой бросок, используя зрительную обратную связь (Messenger, 1968).
Локомоторные ритмы часто генерируются центральными механизмами. Отведение электрической активности моторных и вставочных нейронов у насекомых выявило центральные регулирующие ритмы при ходьбе (например, Pearson, Iles, 1970; Burrows, Horridge, 1974), плавании (например, Kennedy, 1976) и полете (например, Wilson, 1968). Однако на точную структуру поведения часто влияют рефлексы и обратная связь с периферии. Так, устранение проприоцептивных сигналов снижает частоту взмахов крыльев у летящей саранчи, а действие ветра усиливает разряд мотонейрона (Wilson, 1968). По-видимому, периферические стимулы действуют на быструю ходьбу таракана меньше, чем на медленную ходьбу его родичей палочников (Carausius) (Wendler, 1966).
Периферическое управление координацией происходит посредством органов чувств, располагающихся в мышцах и

| Рис. 14.2. Каракатица, хватающая креветку. |

|
| Рис. 14.3. Центральная и периферическая регуляция движения. Движение глазного яблока (вверху) управляется центральной нервной системой. В норме глазное яблоко не подвергается нагрузке, поэтому подробные моторные команды, идущие от мозга, выполняются быстро и точно. Если же на глазное яблоко создать нагрузку (например, надавить пальцем), то его смещение не корректируется. Периферический контроль важен при движении конечностей (внизу), потому что они часто бывают нагружены. Изменение в положении конечности регистрируется сенсорными рецепторами, и по обратной связи о нем сообщается управляющему центру, который корректирует смещение. |
других частях тела; эти органы чувств посылают информацию в мозг и тем самым влияют на его команды мышцам (рис. 14.3). Периферическая регуляция обычно осуществляется совместно с центральной. Например, при координации плавательных движений у рыб головной мозг посылает ритмические сигналы, которые волнами проходят по спинному мозгу, координируя ритмические движения плавников и хвоста. У кошачьей акулы (Scyliorhinus) при перерезке всех нервов, ведущих от мышц к головному мозгу, этот ритм исчезает, но если сохранить некоторые нервы, он продолжается. Следовательно, некоторая периферическая обратная связь, по-видимому, необходима для запуска центрального ритма (Gray, 1950). У хрящевых рыб (Chondrichthyes), в том числе у кошачьей акулы и у всех других акул и скатов, плавники производят небольшие независимые ритмические движения, но у костистых рыб (Teleostei) они при некоторых обстоятельствах ударяют с разной частотой. Хольст (Hoist, 1939, 1973) показал, что ритмы разных плавников могут влиять друг на друга,-это свойство он назвал «относительной координацией». Иногда ритм одного плавника подстраивает под себя ритм другого и доминирует над ним так, что движения происходят в фазу. В других случаях амплитуды движений плавников суммируются, уменьшаясь, когда плавники движутся со сдвигом по фазе и увеличиваясь при совпадении фаз.
У рыб и амфибий локомоция, по-видимому, находится под контролем главным образом эндогенных спинальных ритмов. Нет данных о специальном моторном контроле со стороны переднего мозга. Его удаление у рыб не меняет их позы или локомоции (Bernstein, 1970). У лягушек и жаб это вызывает общее снижение спонтанных движений, но электрическая стимуляция их переднего мозга не приводит к специфическим моторным эффектам. Средний мозг играет некоторую роль в
моторной регуляции у рыб и амфибий. Как было указано в гл. 13. электрическая стимуляция тектума у жаб вызывает поворот головы, а также схватывание и проглатывание корма. У эволюционно более продвинутых позвоночных высший отдел головного мозга в большей степени способен контролировать движения, но автоматические компоненты локомоции по-прежнему регулируются прежде всего его стволом и спинным мозгом.
У млекопитающих кортикоспинальный тракт является самым важным проводящим путем, участвующим в произвольной регуляции движения. Он начинается в двигательной коре и проходит через средний мозг и мозговой ствол к спинному мозгу. Эта система, иногда называемая пирамидной, имеется у всех млекопитающих, за исключением очень примитивных однопроходных (например, утконоса и австралийской ехидны). У сумчатого кистехвостого кускуса (Trichosaunis) пирамидные аксоны доходят только до грудного отдела, где они иннервируют передние конечности. Задние конечности иннервируются экстрапирамидной системой.
Экстрапирамидная система включает все двигательные пути, не входящие в кортикоспинальную или пирамидную системы. Она считается более примитивной системой, чем пирамидная. У животных со слаборазвитой корой больших полушарий или вовсе лишенных ее базальные ганглии экстрапирамидной системы являются основными центрами моторного контроля. Они особенно хорошо развиты у птиц, у которых практически нет коры, но стриатум крупнее, чем у млекопитающих. Таким образом, в ходе эволюции кора больших полушарий, по-видимому, стала вторым источником двигательной координации, действующим через пирамидную систему.
У обезьян нейроны в разных частях кортикоспинальных путей меняют характер своей импульсации при произвольных • движениях глаз, передних и задних конечностей (Evarts. 1968). Искусственная стимуляция двигательной коры вызывает реакции отдельных мышц или отдельных моторных единиц в мышце. Более обширная стимуляция приводит к отдельным

|
| Рис. 14.4. Головной мозг голубя с крупным мозжечком |
движениям целой конечности. Можно нанести карту на поверхность коры в соответствии с теми частями тела, которые движутся в ответ на ее электрическую стимуляцию. Можно построить такую же карту для сенсорной коры, на которой разные части тела представлены по-разному в соответствии с их сенсорным значением для данного вида. Произвольные выученные движения вызываются корковыми пирамидными нейронами, а рефлекторное поддержание двигательных реакций и позы управляются расположенными поблизости экстрапирамидными нейронами. Число пирамидных нейронов у человека оценивается в один миллион (Prosser, 1973).
Другой важной для координации частью головного мозга позвоночных является мозжечок (рис. 14.4). Он участвует в позных рефлексах или в управлении движением косвенно, являясь контролером и координатором нервной активности при ориентации, сохранении равновесия и других тонких реакциях, связанных с регуляцией движения. Основная нейронная организация мозжечка одинакова во всех классах позвоночных - он претерпел меньше эволюционных изменений, чем любая другая часть головного мозга. Мозжечок получает информацию от органов зрения,
слуха, осязания, равновесия, о состоянии мышц и суставов. Он связан также с двигательными областями коры большого мозга. Кроме связей через таламус с двигательной корой он имеет двусторонние связи с сенсорными областями коры, которые играют важную роль в поддержании позы. Вертикальная поза не может сохраняться без зрительной, вестибулярной и проприоцептивной информации. Мозжечок сочетает зрительную и вестибулярную информацию о равновесии с информацией о сокращении соответствующих мышечных веретен и посылает необходимые команды мышцам, в особенности по эфферентным гамма-волокнам (см. гл. 11). Таким образом, мозжечок осуществляет модулирующий тонкий контроль над мышечными сокращениями, связанными с поддержанием позы и со сложными координационными движениями, которые требуются для локомоции.
Пространственная ориентация
Ориентация всего тела животного в пространстве может быть основана на очень простых принципах, но иногда включает и весьма сложные механизмы. Эти простые принципы легче всего наблюдать на некоторых видах беспозвоночных. Френкель и Ганн (Fraenkel, Gunn, 1940) предложили классификацию, основанную на работе более ранних авторов и послужившую отправным пунктом для последующих обсуждений и обзоров (например, Adler, 1971; Kennedy, 1945; Hinde, 1970).
Самая простая форма пространственной ориентации - это кинез, при котором реакция животного пропорциональна интенсивности стимула, но не зависит от его пространственных свойств. Например, мокрицы (Porcellio scaber) стремятся скапливаться во влажных местах под камнями или упавшим деревом. Они активно движутся при низкой влажности и менее активно - при высокой. В результате мокрицы проводят больше времени во влажной среде, и их высокая активность в сухих местах повышает их шансы попасть во влажные условия. Сходное поведение наблюдается у пескоройки - личинки речных миног, активность плавания у которой изменяется в зависимости от интенсивности освещения (Jones, 1955).
Тип кинеза, при котором скорость локомоции связана с интенсивностью стимуляции, называется ортокинезом. Другой тип, наблюдаемый у плоского червя Dendrocoelum lacteum, называется клинокинезом. При нем по мере усиления освещенности меняется скорость изменения направления (Ullyott, 1936; Fraenkel, Gunn, 1940; Hinde, 1970).
При многих типах ориентации, обычно объединяемых в группу таксисов, животное направляется прямо к источнику стимуляции или прямо от него. Например, когда личинка комнатной мухи (Musca domestica) прекращает питаться, она ищет темное место для окукливания. При этом она уползает прочь от источника света, что называют «отрицательным фототаксисом». У личинки на голове имеются примитивные глаза, регистрирующие изменения освещенности, но не способные давать информацию о направлении, в котором находится источник света. Когда личинка ползет, она поворачивает голову из стороны в сторону (рис. 14.5). Если свет слева ярче, чем справа, менее вероятно, что личинка повернет голову влево. Таким образом, она скорее поползет вправо, т. е. подальше от источника света. В ответ на повышение освещенности личинка усиливает скорость поворотов головы. Если свет над головой выключать каждый раз, когда она поворачивает голову вправо, и включать каждый раз при повороте ее влево, тогда личинка будет отворачиваться от освещаемой стороны, двигаясь по кругу вправо. Таким образом, хотя у животного нет дирекциональных рецепторов, оно способно к дирекциональной реакции. Такое же поведение наблюдается у одноклеточного эвглены с одним фоточувствительным «глазком» (Fraenkel, Gunn, 1940).
Ориентация путем последовательного сравнения интенсивности стимула требует поворотов. Обычно она называется клинотаксисом. У многих животных он проявляется в ответ на градиенты химических стимулов. Одновременное сравнение интенсивности стимулов, получаемых двумя

| 
|
| Рис. 14.5. Клинотаксис у личинки мухи. Обратите внимание на движение головы из стороны в сторону. При перемещении находящегося сзади источника света в сторону личинка отворачивается от него. (По Mast, 1911.) | Рис. 14.6. Телотаксис: траектории движения крабов-отшельников при предъявлении им двух источников света (С1 и С2). Каждая часть пути направлена только к одному источнику. (По Fraenkel, Gunn, 1940.) |
или более рецепторами, дает возможность животному выровнять ее. В результате оно переходит к тропотаксису, который позволяет двигаться прямо к источнику стимуляции или от него. Например, мокрица Armadillidium vulgare, живущая под камнями или упавшими деревьями. проявляет положительный фототаксис после периодов высыхания или голода. Благодаря двум сложным глазам на голове животное способно двигаться прямо к источнику света. Если же один глаз зачернить, оно движется по кругу. Это говорит о том, что в норме оба глаза уравновешивают стимуляцию. При предъявлении двух световых источников мокрица часто начинает с того, что движется в среднем между ними направлении, а затем поворачивает к одному из них. Это происходит потому, что равная стимуляция обоих глаз достигается при движении либо точно между двумя источниками, либо прямо к одному из них. Отклонения от одного источника являются саморегулирующимися, а боковой свет игнорируется, потому что сзади и с боков глаза экранированы. В то же время отклонения от среднего маршрута между двумя широко расставленными источниками могут привести к потере контакта с одним из них.
Животные с глазами, дающими благодаря своему строению информацию о направлении света, способны к телотаксису форме дирекциональной ориентации, которая не зависит от одновременного сравнения стимуляции двух рецепторов. При двух источниках стимуляции животное движется к одному из них и никогда не избирает среднего направления: это показывает, что влияние одного из стимулов тормозится. Такой пример приведен на рис. 14.6.
Менотаксис - этотакая форма телотаксиса (Hinde. 1970), при которой ориен-
| Рис. 14.7. Солнечный компас: муравей (Lasius niger) возвращается в гнездо Г, когда солнце светит справа под углом около 90 к направлению движения животного. В точке X муравей был задержан на 2,5 ч. Когда его выпустили, он отклонился от своего прежнего пути на такой же угол, на какой переместилось за это время солнце, и его лучи снова составили угол в 90° с направлением движения (По Brun, 1914.) | 
|
тация происходит под углом к направлению стимуляции. Примером служит реакция на «световой компас» у возвращающихся в муравейник муравьев. Эти животные отчасти ориентируются по положению солнца. Медленно меняя его видимое положение при помощи зеркала, можно заставить муравьев соответственно изменить свой путь (рис. 14.7). Раньше полагали (Brun, 1914), что если муравьев (Lasius niger), возвращающихся домой, посадить на несколько часов в темный ящик, то при освобождении они сохранят тот же угол движения по отношению к солнцу, что и до пребывания в ящике. Но затем стало очевидным (Jander, 1957), что муравьи делают поправку на перемещение солнца и по освобождении движутся в том же направлении. Такие компенсированные во времени компасные реакции продемонстрированы также у жука-навозника Geotrupes sylvaticus, прудовой водомерки Velia currens имедоносной пчелы Apis mellifera (см. обзор Saunders, 1976).
Тип ориентации, осуществляемый в этом случае, зависит как от природы внешних сигналов, так и от сенсорного оснащения организма. Животное, воспринимающее лишь силу стимула, ограничивается ее последовательными измерениями в разных пунктах. Если внешние сигналы по своей природе имеют направленность, единственный односторонне экранированный рецептор может предоставить дирекциональную информацию. Экранированный фоторецептор в этом отношении полезен, но экранированный хеморецептор никаких преимуществ не дает, потому что химические стимулы по природе своей не дирекциональны. При двух рецепторах одновременное сравнение может быть использовано для обнаружения градиентов (рис. 14.8). Если множество рецепторов расположено в форме растра (т.е. ряда или мозаики), возможны более сложные типы ориентации (рис. 14.8). Примерами растров служат глаза с хрусталиком у позвоночных и сложные глаза членистоногих (см. гл. 12). Пространственная ориентация часто достигается сочетанием разных методов. Например, самцы некоторых бабочек привлекаются самками с помощью испускаемого самкой и распространяющегося по воздуху феромона. Этот запах разносится ветром, поэтому летящий самец должен учитывать движение воздуха. Бабочки в полете обычно пользуются зрительными ориентирами, чтобы контролировать свое продвижение относительно почвы. На маршрут животного в воздухе влияет направление ветра, что и определяет его окончательный путь (рис. 14.9). Результаты опытов с бабочками показывают, что угол полета меняется с концентрацией запаха. В отсутствие запаха животное ле-
тает взад и вперед, не продвигаясь против ветра (т. е. под прямым углом к нему). Когда ветер приносит запах, угол полета увеличивается и животное движется зигзагами против ветра. Изменения направления связаны с границами запахового следа (рис. 14.9). При снижении концентрации запаха ниже определенного уровня, в частности у края его струи, животное начинает двигаться в противоположном направлении. Эти повороты не связаны с направлением ветра, а зависят от внутреннего эталона, или идиотетической информации. Таким образом в поисках полового партнера летящая бабочка использует сочетание зрительных, анемотаксических (связанных с ветром) и идеотетических ориентационных механизмов.
Принцип реафферентации
Высокоразвитая система ориентации должна быть способна отличать стимулы из внешнего мира от стимуляции, создаваемой самим животным. Так, например, что касается зрения, то у человека перемещение объектов во внешнем мире вызывает движение изображения по сетчатке, которое мы воспринимаем. Однако произвольное движение глаз тоже вызывает перемещение изображения на сетчатке, но оно уже не воспринимается. Каким-то способом мозг отличает перемещение изображения по сетчатке, независимое от животного, от перемещения, вызываемого движением самого глазного яблока.
Для объяснения этого явления предложены две теории - оттока и притока. Согласно теории оттока, основы которой заложил Гельмгольц (Helmholtz, 1867). команды глазным мышцам о движении глазного яблока сопровождаются параллельными сигналами, идущими к компаратору в головном мозгу. Здесь они сравниваются с приходящими зрительными сигналами (рис. 14.10). Согласно теории притока, созданной Шеррингтоном (Sherrington, 1918), рецепторы наружных глазных мышц посылают сообщения в мозговой компаратор при любых движениях глаз (рис. 14.10). В обеих теориях компаратор оценивает оба приходящих

| Рис. 14.8. Схематическое изображение некоторых основных принципов сенсорной ориентации. А. Направление стимуляции (например, снега) регистрируется растром сенсорных рецепторов. Б. Направление регистрируется посредством сравнения одновременной стимуляции двух рецепторов. В. Имеется только один рецептор, и животное проводит последовательные сравнения, передвигая туловище. Г. Время прихода стимула (например, "звуковых волн) сравнивается двумя рецепторами. Д. Градиент стимуляции (например, химической) peгистрируется посредством сравнения данных, получаемых двумя рецепторами. Е. Градиент регистрируется одним рецептором, когда животное движется, обследуя разные точки пространства. (Из The Oxford Companion to Animal Behaviour, 1981.) |
сигнала и определяет, соответствуют ли зрительные сигналы тому движению, которое следовало ожидать на основании второго сигнала. Если сигналы не соответствуют друг другу, значит, какая-то часть движения должна была быть вызвана внешними причинами.
Наружные глазные мышцы содержат

|
| Рис. 14.9.Самец бабочки, летящей против ветра на феромон, выделяемый самкой. На направление полета П влияет направление ветра В, и возникает результирующий маршрут Р. Угол передвижения - это угол между направлением ветра В и направлением Р по отношению к земле. |
мышечные веретена, и их существование, казалось бы, подтверждает теорию Шеррингтона. Однако, по-видимому, эти мышцы не участвуют в подаче сигналов о положении глаза (Merton, 1964; Howard, Templeton. 1966). Для доказательства того, что в глазных мышцах нет рецепторов положения, Гельмгольц (Helmholtz, 1867) пользовался данными по механическим манипуляциям с глазным яблоком и кажущимся движениям, вызываемым попытками двигать глазом, когда наружные глазные мышцы парализованы.
Как хорошо известно, при смещении глазного яблока в глазнице нажимом на него пальцев зрительная ось сдвигается (что можно заметить по раздвоенному изображению); она остается сдвинутой, пока длится нажим. Глазное яблоко не давит на палец чтобы занять свое прежнее положение, как можно было бы ожидать, если бы в регуляции положения глаза участвовали мышечные веретена (McFarland, 1971). Кроме того, при смещении глазного яблока пальцем движение ощущается, чего не должно бы быть по теории притока. Мышечные веретена возбуждались бы при любом движении глазного яблока, и тогда компаратор в мозгу компенсировал бы движение изображения по сетчатке. Поэтому факты, по-видимому, говорят в пользу теории оттока.

| Рис. 14.10. Теории притока и оттока сигналов (импульсов) при управлении движением глазного яблока. |
Теория оттока была развита и обобцена Хольстом и Миттельштедтом Holst, Mittelstaedt. 1950; см. также Holst, 1954). Согласно их принципу реафферентации, мозг отличает экзафферентную стимуляцию (вызываемую только факторами вне животного) от реафферентной, происходящей в результате движений тела животного). Моторные команды не только организуют мышечные движения, но и создают их нервную копию (копию "эфферентации»), соответствующую сенсорным сигналам, которые можно было ожидать при данном поведении животного. В результате мозг сравнивает копию эфферентации с приходящей сенсорной информацией (рис. 14.11). Вся реафферентная информация должна быть погашена копией эфферентации так, чтобы выход из компаратора был равен нулю. В то же время экзафферентная информация нe аннулируется, а передается компаратором в другую часть мозга.
Хольст и Миттельштедт (Holst, Mittelstaedt, 1950), показали, что если двукрылую ильницу (Eristalis) поместить в цилиндр с вертикальными полосами, у нее наблюдается типичный оптомоторный рефлекс, т. е. она поворачивается в направлении полос при вращении цилиндра. Такие рефлексы не возникают, если ильница движется сама по себе, хотя зрительная стимуляция при этом такая же. Если в опыте повернуть этому насекомому голову на 180 (рис. 14.12), то, как и следовало ожидать, оптомоторный рефлекс становится обратным. Но если ильница пытается двигаться сама, го начинает вертеться на месте, и ее движения, по-видимому, самовозбуждаются. Эти данные можно объяснить, исходя из теории реафферентации. В норме выходные сигналы компаратора определяют движение тела, а когда двукрылое движется само по себе, выход равен нулю и движения нет. Оптомоторный аппарат выдает экзафферентную стимуляцию, которая не аннулируется копией эфферентации, и ильница отвечает рефлексом. Когда голова ее повернута на 180 , экзафферентная стимуляция вызывает то же действие, что и раньше, но в обратном направлении. Однако в случае реафферентной стимуляции воспринимаемое движение имеет обратный знак и не вычитается из копии эфферентации, а прибавляется к ней, и в

|
| Рис. 14.11. Схема основной системы реафферентации. |
результате выход компаратора, который в норме должен быть сведен к нулю, возрастает. Чем больше животное реагирует, тем сильнее реафферентная стимуляция и тем больше усиливается реакция животного. В результате насекомое крутится на месте все быстрее и быстрее. Принцип реафферентации важен не только в отношении зрения, но также для контроля за положением конечностей, тела и т.д. Так, например, мы можем отличить движения руки, трясущей ветку дерева, от, возможно, таких же движений, возникающих, когда рука пассивно держится за ветку, раскачиваемую ветром.
Навигация
Самая сложная форма пространственной ориентации - это навигация. Она требует не только компаса, или дирекционального чувства, но и своего рода карты. Для иллюстрации рассмотрим опыт, проведенный Бейкером (Baker, 1981) для проверки того, может ли человек пользоваться магнитной информацией. Группа студентов была перевезена в небольшом автобусе от определенной начальной точки (дома) к некоторому секретному месту назначения. По приезде студенты должны были показать, в какой стороне находится их дом. Для точного ответа необходим внутренний магнитный компас, и Бейкер искал данные в пользу его наличия. Однако выполнение такой задачи требует также знания относительного положения обоих пунктов. Даже если бы студенты обладали точным магнитным компасом, они не могли бы указать, в каком направлении находится дом, без какой-нибудь карты.
При навигации важны три типа ориентации: 1) пилотирование, или прокладывание пути по знакомым ориентирам; 2) компасная ориентация, т. е. способность двигаться по определенному азимуту, не пользуясь ориентирами, и 3) истинная навигация, или способность направляться к цели - дому или месту выведения потомства, — не пользуясь ни ориентирами, ни компасом. В своих дальних миграциях птицы, вероятно, используют все три типа ориентации. Пердек (Perdeck,

|
| Рис. 14.12. Ильница Eristalis во вращающемся цилиндре. А. Нормальное положение головы. Б. Голова повернута на 180°. Л - левый глаз; П - правый глаз. (По van Hoist, 1954.) |

|
| Рис. 14.13. А. Отлов размножавшихся и зимовавших скворцов, окольцованных в Нидерландах. Б. Отлов взрослых и молодых скворцов, перевезенных из Нидерландов в Швейцарию и выпущенных там во время осенней миграции. (По Perdeck, 1958.) |
1958, 1967) отлавливал молодых и взрослых скворцов, когда они пролетали через Нидерланды при своей первой осенней миграции, кольцевал их и перевозил на самолете в Швейцарию, т. е. на 750 км юго-восточнее их обычного миграционного пути. В норме скворцы мигрируют от своих гнездовий на Балтике к зимовьям в Бельгии, южной Англии и северной Франции (рис. 14.13). После перемещения в Швейцарию молодых скворцов ловили в Испании и южной Франции (рис. 14.13); это говорит о том, что они сохраняли нормальное юго-западное направление миграции. Но взрослых скворцов снова ловили в местах их обычных зимовок. Таким образом, взрослые скворцы внесли поправку на свое перемещение, тогда как молодые сохранили компасную ориентацию, характерную для их местной популяции. Молодые животные многих мигрирующих видов в первый раз достигают мест своих зимовок на основе врожденной информации о направлении на цель и расстоянии до нее. Они не способны внести поправку на перемещение, потому что у них нет карты, необходимой для истинной навигации (Schmidt-Koenig, 1979).
Ориентация по окружающим объектам может играть важную роль в навигации, особенно при приближении к месту назначения. Классический пример этого приведен в работе А. Хаслера (A. Hasler, 1960) по миграции лосося. Тихоокеанские лососи (Oncorhynchus) вылупляются из икры в ручьях западной части США и Канады. Проходя стадию смолта, они скатываются вниз по реке в Тихий океан. Проведя два-три года в море, лососи становятся половозрелыми и возвращаются в тот же самый ручей, где родились. Путешествие к берегу, вероятно, происходит по солнечному компасу. Но дойдя до берега, они должны выбрать нужную реку и единственный из впадающих в нее ручьев. После многих лет работы Хаслер с сотрудниками установили, что на стадии смолта в рыбах «запечатлеваются» запаховые признаки их родного ручья (импринтинг). Во время обратного путешествия они отличают его воду от воды других речных притоков. В сущности они опознают ориентиры, но, поскольку запах

|
| Рис. 14.14. Прибор для выработки условного сердечного рефлекса у голубей при испытаниях их чувствительности к разным стимулам (в данном случае - к поляризованному свету). (По SchmidtKoenig, 1979.) |
места их рождения распространяется водой по всему пути миграции, молодому лососю не нужно запоминать его по дороге к океану. И действительно, если после периода импринтинга лососей перевести в другой ручей, они вернутся в свою родную речку, а не в ту, по которой мигрировали к океану.
Известно, что животные пользуются разного рода компасами, основанными на свойствах геофизической среды, например магнитным полем земли. Доказать, что определенное физическое свойство используется в качестве компаса, можно, продемонстрировав способность животного обнаруживать это явление и использовать его для ориентации в естественных условиях. Ввиду неуправляемой изменчивости природы проверять сенсорные способности лучше всего в лаборатории. Излюбленным методом исследователей навигации служит метод условных сердечных рефлексов (рис. 14.14). Этим и другими методами было показано, что голуби чувствительны к следующим стимулам:
Внешнее давление. Голубь чувствителен к изменениям атмосферного давления в пределах от 1 до 10 мм водяного столба, что соответствует изменению высоты менее 10 м (Kreithen, Keeton, 1974; Delius, Emmerton, 1978); благодаря такой сенсорной способности голубь обладает точным физиологическим альтиметром.
Инфразвук. Звук частотой менее 10 Гц называется инфразвуком. Человек не может слышать его, голуби же воспринимают частоты до 0,06 Гц. Инфразвук распространяется на очень большое расстояние, и такие его источники, как прибой, голуби могли бы использовать для навигации (Yodlowski et al., 1977; Kreithen, 1978).
Запах. Долго считалось, что у птиц слабое обоняние, но опыты с условными сердечными рефлексами (например, Henton et al., 1966; Shumake et al., 1966) подтверждают результаты применения физиологических методов и показывают, что обоняние у голубей достаточно тонкое, чтобы служить при навигации. Такие же данные получены для других видов птиц (Schmidt-Koenig, 1979).
Магнитный компас. Ученые долго считали, что энергия геомагнитных явлений слишком мала для того, чтобы животные могли ее воспринимать. Теперь известно, что это не так, и у многих видов показаны реакции на магнитные поля.
Хотя и раньше были указания на магнитную чувствительность у птиц (например, Merkel, Wiltschko, 1965), она долго вызывала сомнения, поскольку ее не удавалось показать в опытах с условными рефлексами на сердце. Но затем были получены положительные результаты не на привязанных, а на свободно передвигающихся голубях (Bookman, 1978).
Миниатюрные магниты обнаружены у бактерий, пчел и голубей. Хотя известно, что пчелы и голуби чувствительны к магнитным полям, не ясно, как магнитная информация воспринимается нервной системой. Имеются данные о том, что маг-

|
| Рис. 14.15. Результаты опытов на почтовых голубях со смешением времени суток. Птиц в круглой клетке обучали искать пишу в определенном азимутальном направлении. Учитывалось число клеваний кормушек, помещенных по окружности (каждая точка - один удар). ЦЕВ- центральноевропейское время. Стрелки - ожидаемое в условиях опыта направление. Верхний ряд - опыты. нижний одновременные контроли. (По Schmidt-Koenig, 1979.) |
нитные явления влияют на биологические часы пчел (Gould. 1980). У птиц чувство направления, по-видимому, связано со склонением магнитного поля Земли, и имеется предположение, что оно же важно для чувства карты у почтовых голубей (см. ниже).
Солнечный компас. Способность птиц использовать солнце как компас открыта Крамером (Kramer, 1951). Он приучал скворцов, находящихся в круглой клетке, искать корм в определенном азимутальном направлении. Все зрительные ориентиры были исключены, и видны были только солнце и небо. Птицы были способны находить нужный азимут в течение всего дня, что доказывает их способность делать поправку на движение солнца.
Биологические часы скворца (см. гл. 16.3) можно искусственно «перевести» (Hoffman. 1954), поместив птицу в светонепроницаемое помещение с искусственным фотопериодом. Шмидт-Кёниг (Schmidt-Koenig. 1958, 1960. 1961) в опытах с почтовыми голубями испытывал действие такого перевода на 6 ч вперед и на 6 или 12 ч назад. Результаты, представленные на рис. 14.15. говорят о том, что ориентация птицы в экспериментальном аппарате была точно такой же, какую можно было ожидать, пользуясь в качестве компаса солнцем.
Показано, что солнечный компас голубей достаточно точен для навигационных целей при условии, что в течение путешествия птицы время от времени делают поправочные измерения. Однако связь азимута солнца с местным временем пригодна прежде всего для опреде-, ления долготы, а с изменениями широты местонахождения наблюдателя меняется высота солнца над горизонтом. По некоторым данным опытов с инструментальными условными рефлексами, голуби могут достаточно точно определять изменения этой высоты. Имеются также данные, что они оценивают высоту солнца, измеряя скорее тени, а не само положение светила (McDonald, 1972, 1973). Действительно, размеры теней могут изменяться в шесть раз сильнее, чем высота солнца над горизонтом, но неизвестно, способны ли голуби использовать эту информацию вне помещения. Уайтен (Whiten, 1972) обучал голубей связывать высоту солнца с северным или южным на-
правлением от дома. Его опыты показывают, что голуби способны сопоставлять эту высоту с прохождением меридиана, однако нет данных об использовании такой способности при навигации.
Некоторые исследователи (например, Matthews, 1955, 1968; Pennycuick, I960) пробовали объяснить навигацию голубей, исходя из движения солнца. Теоретически информации достаточно для определения долготы (азимут солнца с поправкой на время) и широты (высота солнца над горизонтом в разное время), т. е. для построения эквивалента карты. Но это требует очень большой остроты зрения и точности увязывания движения солнца с временем. Большинство ученых сомневается в том, что голуби способны на такую точность (Schmidt-Koenig, 1979). Кроме того, способность летящего голубя определять высоту солнца никогда не была доказана, а между тем это весьма важно с точки зрения теорий навигации по солнцу.
Звездный компас. Если певчие птицы посажены в клетку в то время, когда обычно происходит их миграция, то они проявляют типичное дирекциональное миграционное беспокойство; ночью эта дирекциональность связана со звездами (Sauer, Sauer, 1955). Птицы были ориентированы под естественным звездным небом и под небом планетария. Методики регистрации миграционного беспокойства позднее были усовершенствованы (Emlen, Emlen 1966) с помощью аппарата, показанного на рис. 14.16. Эмлен (Emlen, 1967) обнаружил, что дирекциональность у индиговых овсянок (Passerina суапеа) одна и та же под естественным небом и под стационарным небом планетария. В то же время птицы следовали за смещением планетарного неба, и Эмлен (1972) обнаружил, что их ориентация была связана скорее с его вращением, чем с определенными созвездиями.
Наземному наблюдателю кажется, что звезды расположены на внутренней поверхности сферы, называемой небесной сферой. В каждый данный момент звезды расположены на небесной сфере определенным образом, причем образуемый ими рисунок движется так, что создает впечатление, будто эта сфера вращается. В действительности вращается Земля вокруг своей оси, проходящей через полюсы. Точка в северном небе, вокруг которой небесная сфера будто бы вращается, называется северным полюсом мира. Полярная звезда лежит очень близко к этому полюсу, и для практических целей ею можно пользоваться при определении направления на север от наблюдателя.
Эмлен (1972) показал, что овсянки, выращенные в неволе и совсем не видевшие неба, не могли ориентироваться во время миграции. Оперившиеся птенцы, помещенные до наступления осенней миграции под небо планетария, вращающееся вокруг Полярной звезды, показывали нормальную ориентацию на юг, А на север ориентировались птицы, находившиеся под экспериментальным небом планетария, которое вращалось вокруг звезды Бетельгейзе в созвездии Ориона, видимой наблюдателю в северных широтах в южной части неба. По-видимому, только что оперившиеся птенцы научались узнавать, какая часть неба вращалась меньше всего, и во время миграционного беспокойства ориентировались в противоположном направлении.
Методики условных сердечных рефлексов тоже были использованы для исследования восприятия птицами ночного неба. Получены данные, что кряквы (Anas platyrhynchos) научаются распознавать определенные расположения звезд (Wallraff, 1969). Хотя такие картины потенциально сходны по своей информации с картой, имеющиеся пока сведения говорят о том, что звезды используются только как компас (Schmidt-Koenig, 1979).
Поляризованный свет. В нормальном неполяризованном свете световые волны колеблются одинаково во всех плоскостях. В поляризованном свете более сильные колебания распространяются в одной плоскости. При рассеянии неполяризованного солнечного света молекулами атмосферы происходит поляризация, максимальная для света, рассеянного под углом в 90° к солнечным лучам, а это значит, что поляризация распределена в небе определенным образом, причем ее распределение меняется в соответствии с положением солнца.
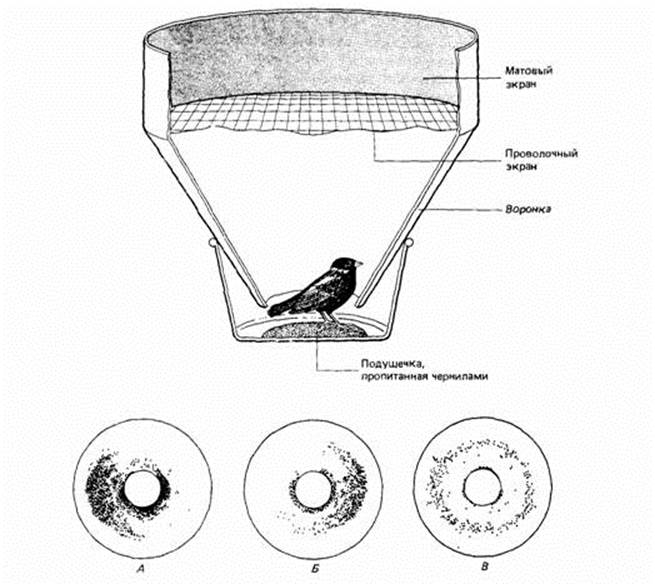
|
| Рис. 14.16. Экспериментальная клетка для измерения миграционного беспокойства. Пытаясь покинуть клетку, птица оставляет чернильные следы на промокательной бумаге, выстилающей воронку. Внизу - несколько примеров регистрации. (По Schmidt-Koenig, 1979.) |
Было обнаружено (von Frisch, 1967), что ориентация пчел, выражаемая их танцем (см. гл. 23), зависит от положения солнца, даже когда оно скрыто тучами. Оказывается, пчелам достаточно видеть только небольшую часть неба, причем нужная информация соответствует ультрафиолетовой части спектра. При пропускании ультрафиолетового света солнца через поляризующий фильтр ориентация танца пчелы менялась в соответствии с углом поляризации.
Опыты с условными сердечными рефлексами (см. рис. 14.14) ясно показывают, что голуби способны воспринимать вращение плоскости поляризации света (Kreithen, Keeton, 1974; Delius et al., 1976), но неизвестно, как они интерпретируют эту информацию. Хотя характер поляризации солнечного света может дать сведения о местоположении солнца даже тогда, когда небо покрыто тучами, по всей вероятности, голуби не пользуются своим солнечным компасом при таких условиях (Schmidt-Koenig, 1979).
ДЛЯ ЗАПОМИНАНИЯ
1. Координация движения подчиняется как центральному, так и периферическому управлению. Периферическое управление преимущественно задано заранее, тогда как центральное может включать в себя обратную связь (замкнутая петля) или же быть предсказующим (незамкнутая петля).
2. Пространственная ориентация простых животных основана на кинезах и таксисах, типы которых зависят от сенсорных способностей животного. Навигация как более сложная форма ориентации требует эквивалентов как карты, так и компаса.
3. Известно, что в навигации участвует множество разных сенсорных модальностей, но не совсем понятно, как координируется информация.
Рекомендуемая литература
Fraenkel G.S., Gunn D.L. (1940) The Orientation of Animals, Clarendon, Oxford (Dover Books, 1961).
Schmidt-Koenig K. (1979) Avian Orientation and Navigation, Academic Press, London.
Гомеостаз и поведение
Понятие физиологической стабильности неотделимо от концепции Клода Бернара (Claude Bernard. 1859) о внутренней среде. Он установил, что уровень сахара в крови остается постоянным независимо от того, голодало животное, только что получало мясо или потребляло корм, содержащий сахар. Он постулировал наличие некоторого процесса регуляции и контроля для поддержания постоянства внутренней среды. Он понимал также, что животное, способное регулировать свою внутреннюю среду при колебаниях внешней среды, способно использовать большее разнообразие потенциальных местообитаний.
Животных можно приблизительно разделить на конформеров, допускающих влияние на свою внутреннюю среду внешних факторов, и на регуляторов, которые удерживают ее в состоянии, в значительной степени независимом от внешних условий. Процессы, посредством которых регуляторы управляют своим внутренним состоянием, объединяются термином гомеостаз.
Гомеостаз
Этот термин впервые был применен американским физиологом Кэнноном (Cannon. 1932), который писал: «Координированные физиологические процессы, которые поддерживают большую часть устойчивых состояний в организме, так сложны и настолько специфичны для живых существ, поскольку могут включать совместную работу головного мозга и нервов, сердца, легких, почек и селезенки, - что я предложил специальное название для этих состояний, а именно гомеостаз». Кэннон рассмотрел ситуацию, в которой сенсорные процессы, следящие за внутренним состоянием организма, вызывают соответствующее действие, как только внутреннее состояние отклонялось от заданного или оптимального. Так, например, когда температура человеческого тела поднимается выше 37 С, вступают в действие такие охлаждающие механизмы, как прилив крови к коже и потоотделение. Когда температура падает ниже оптимального уровня, включаются согревающие механизмы, например дрожь. Пользуясь множеством таких тонко настроенных механизмов, человек достигает точной терморегуляции и теплового гомеостаза.
Хотя типы регуляторных механизмов, рассмотренные Кэнноном, как теперь известно, широко распространены в животном мире и включают самые разнообразные физиологические процессы, в регуляции внутренней среды участвуют не только они. Так, например, до недавнего времени считали, что животные усиленно пьют при высокой окружающей температуре в результате обезвоживания, которое возникает при таких охлаждающих реакциях, как потоотделение и одышка. Эта точка зрения полностью соответствует изложенной выше теории гомеостаза: испарение воды необходимо для поддержания теплового гомеостаза в жаркой среде, а это нарушает в организме водный баланс, восстановление которого требует усиленного питья. Однако теперь мы знаем, что у таких видов, как крыса и голубь, питье прямая реакция на температурное изменение, опережающая любое нарушение водного баланса в результате терморегуляции, а не ответ на нарушение.
как считает регуляторная теория (Budgell, 1970а, b). Иными словами, животные пьют, чтобы запастись водой для терморегуляции.
Роль поведения в регуляции внутренней среды значительно отличается у разных видов и зависит от обстоятельств. Питье, например, нужно для поддержания гомеостаза у многих видов, так как физиологические механизмы не могут за
|
Просмотров 688 |
|
|
