Главная Обратная связь
Дисциплины:
Архитектура (936)
Биология (6393)
География (744)
История (25)
Компьютеры (1497)
Кулинария (2184)
Культура (3938)
Литература (5778)
Математика (5918)
Медицина (9278)
Механика (2776)
Образование (13883)
Политика (26404)
Правоведение (321)
Психология (56518)
Религия (1833)
Социология (23400)
Спорт (2350)
Строительство (17942)
Технология (5741)
Транспорт (14634)
Физика (1043)
Философия (440)
Финансы (17336)
Химия (4931)
Экология (6055)
Экономика (9200)
Электроника (7621)

ПРОДОЛГОВАТЫЙ МОЗГ И ВАРОЛИЕВ МОСТ
|
|
Продолговатый мозг и варолиев мост вместе составляют задний мозг. Анатомически и функционально задний мозг связан со спинным мозгом, мозжечком и большими полушариями головного мозга. В нем замыкаются дуги многих сложнокоординирован-ных двигательных процессов.
Сегментарное строение, четко выраженное в спинном мозге, стерто в продолговатом. Сюда вступают афферентные волокна от слуховых рецепторов, рецепторов полости рта, кожи лицевой части головы, сердца, крупных сосудов и легких, слизистых оболочек глаз, ряда рецепторов желудка, печени, поджелудочной железы, тонких кишок. Но в органах грудной и брюшной полости имеются также рецепторы, от которых афферентные волокна идут не в продолговатый, а в спинной мозг. Эфферентные волокна нейронов продолговатого мозга иннервируют все мышцы лица, сердце, бронхи, гортань, пищевод, желудок, поджелудочную железу, печень, кишечник. Иннервация внутренних органов осуществляется парасимпатическими волокнами совместно с симпатическими волокнами, лежащими в спинном мозге.
Из области продолговатого мозга и варолиева моста выходят следующие черепно-мозговые нервы: тройничный, отводящий, лицевой, слуховой, языкоглоточный, блуждающий, добавочный, подъязычный.
Продолговатый мозг играет большую роль в регуляции мышечного тонуса. Скелетные мышцы находятся в постоянном тонусе, за счет которого преодолевается действие силы тяжести и тело сохраняет равновесие. Тонус мышц имеет рефлекторную природу. Начало дуги этого рефлекса лежит в мышцах и сухожилиях.
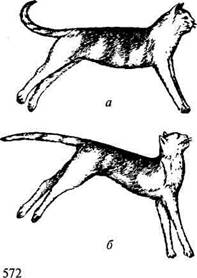
Если головной мозг вместе со средним мозгом и его красными ядрами удален, а продолговатый мозг сохранен и не отделен от спинного, то у млекопитающих развивается сильное, часами и сутками удерживаемое напряжение разгибательных мышц (рис. 13.13).
|
Конечности такого животного вытянуты, и согнуть их можно лишь с трудом; шейные мышцы, разгибающие голову, напряжены; позвоночник несколько выгнут дугой кверху.
Таким образом, все мышцы, напряжение которых противостоит действию силы тяжести, оказываются после отделения продолговатого мозга от среднего в состоянии длительного тонического напряжения. Это
Рис. 13.13. Децеребрационная ригидность:
а — кошка с выключенными большими полушариями (путем перевязки сосудов); б— то же при одновременном выключении мозжечка
резкое усиление тонуса разгибательных мышц носит название децеребрационной ригидности (децеребрация — удаление мозга выше продолговатого). Тонус разгибателей при децеребрационной ригидности превышает тонус сгибателей, но длительное напряжение сгибателей способствует тому, что конечности становятся как бы твердыми опорами и могут удерживать тяжесть тела.
Децеребрационная ригидность не наступает, если средний мозг животного связан с продолговатым мозгом, а также если продолговатый отделен от спинного мозга. Отсюда следует, что в продолговатом мозге лежат нервные клетки, которые могут приводить в возбуждение спинальные нейроны, иннервирующие разгибатели. Если продолговатый мозг связан со средним мозгом, ригидность также не наступает, т. е. от среднего мозга исходят влияния, угнетающие стимуляцию разгибательных рефлексов со стороны продолговатого мозга.
Заключительный этап в возникновении непомерного тонуса связан с рефлекторной деятельностью ядра вестибулярного нерва. Волокна от рецепторов вестибулярного аппарата в продолговатом мозгу контактируют с массой клеток, образующих так называемое вестибулярное ядро. Оперативное удаление вестибулярного ядра и особенно его части, известной как ядро Дейтерса, резко ослабляет децеребрационную ригидность. Разрушение вместе с ядром Дейтерса ретикулярной формации продолговатого мозга совершенно ее прекращает.
Для развития децеребрационной ригидности необходимо поступление в ЦНС импульсов от рецепторов разгибательных мышц. Инъекция в эти мышцы раствора новокаина или перерезка соответствующих задних корешков снимает тонус мышц-разгибателей. Следовательно, децеребрационную ригидность надо рассматривать как результат усиления рефлексов, осуществляемых совместно спинным и продолговатым мозгом в ответ на импульсы, поступающие как от рецепторов мышц-разгибателей, растягиваемых под действием силы тяжести, так и от рецепторов лабиринта. Эти рефлексы в норме всегда уравновешиваются воздействиями, рефлекторно возникающими в среднем мозге, мозжечке, а у высших млекопитающих главным образом в коре больших полушарий.
Роль продолговатого мозга в осуществлении децеребрационной ригидности обусловлена постоянным поступлением импульсов от важнейших рецепторов дыхательной и кровеносной систем, а также с лабиринтов, вследствие этого он постоянно воздействует на рефлекторные центры спинного мозга, повышая их возбудимость. При отделении спинного мозга от продолговатого возбудимость нервного центра никогда не достигает той степени, при которой тонус мышц-разгибателей в ответ на импульсы от их рецепторов рефлекторно усиливался бы до степени, характеризующей децеребрационную ригидность.
Децеребрационная ригидность — это состояние, конечно, совершенно нефизиологичное, но по ней можно судить о взаимоотношениях между разными образованиями ЦНС. Надо заметить, что механизм этого процесса еще не совсем ясен. Это состояние не всегда наступает после децеребрации, иногда оно выражено очень резко, а иногда почти не проявляется.
Продолговатый мозг участвует в регуляции проводниковых рефлексов и в осуществлении таких актов, как ходьба и стояние, для которых необходима интегративная деятельность ЦНС. Буль-барное животное, у которого сохранены лишь спинной и продолговатый мозг, способно к более сложным рефлекторным процессам, чем спинальное, так как все основные функции в более полной мере объединены и скоординированы.
Варолиев мост является центральной частью заднего мозга. Он имеет вид толстого белого вала. Серое вещество образуют ядра черепно-мозговых нервов (V—VIII пары), ядра ретикулярной формации и собственные ядра моста.
Белое вещество состоит из продольных и поперечных пучков нервных волокон. Продольные волокна идут по нижней поверхности моста в продолговатый мозг, образуя в нем пирамиды. Сверху они составляют восходящие и нисходящие пути двусторонних связей моста. Поперечные волокна направляются в ножки мозжечка.
Чувствительные волокна черепно-мозговых нервов проводят сигналы от вкусовых рецепторов языка, рецепторов кожи и мышц головы, глаз и зубов. Эфферентные волокна обеспечивают мимику. Таким образом, рефлекторная деятельность варолиева моста дополняет защитные и пищевые рефлексы, осуществляемые продолговатым мозгом.
СРЕДНИЙ МОЗГ
Средний мозг, так же как продолговатый мозг и варолиев мост, относится к стволовым структурам. Он представляет собой относительно небольшую часть мозгового ствола. Через средний мозг проходят все восходящие пути, передающие импульсы к таламусу, большим полушариям и мозжечку, а также нисходящие пути, проводящие импульсы к продолговатому и спинному мозгу. В среднем мозге, так же как и в продолговатом, расположены нейроны ретикулярной формации.
Анатомически средний мозг состоит из двух основных частей: дорсального отдела, который представляет собой крышку мозга, или четверохолмие, и базального, который представлен его ножками. Каждая из ножек, в свою очередь, состоит из покрышки, черной субстанции и основания. Покрышка — это продолжение дорсальной части моста, в которой находятся красное ядро и ядро блокового и глазодвигательного нервов (рис. 13.14).

|
Рис. 13.14. Разрез через средний мозг на уровне ядер глазодвигательного нерва:
/ — ножки мозга; 2— глазодвигательный нерв; 3— красное ядро; 4— ядро глазодвигательного нерва; 5— центральное серое вещество; б—водопровод мозга (сильвиев водопровод); 7— переднее двухолмие; 8— черная субстанция
Рефлекторные функции ядер среднего моз-г а. Передние бугры четверохолмия являются первичными зрительными центрами, при участии которых осуществляются некоторые рефлексы в ответ на световые раздражения. К таким рефлексам относятся зрительные ориентировочные рефлексы, проявляющиеся в том, что животное, лишенное больших полушарий, но с сохраненным средним мозгом, реагирует на световое раздражение движением глаз и поворотом головы. Рефлекторные движения глаз происходят благодаря поступлению к глазным мышцам импульсов от ядер глазодвигательного и блокового нервов. Передние бугры четверохолмия принимают участие в осуществлении зрачкового рефлекса, аккомодации глаза, а также в конвергенции — сведении зрительных осей.
Задние бугры четверохолмия представляют собой первичные слуховые центры. При их участии осуществляются ориентировочные слуховые рефлексы. При ориентировочных рефлексах у животных с целым средним мозгом наблюдаются и вегетативные рефлексы: изменение ритма сердечной деятельности, артериального давления.
Ядра четверохолмия обеспечивают сторожевой рефлекс, значение которого состоит в подготовке реакции организма на внезапное раздражение. При этом происходит перераспределение мышечного тонуса: усиление тонуса сгибателей, что способствует быстрой ответной реакции животного.
В ножках мозга идут проводящие пути как от нижележащих центров в высшие центры головного мозга, так и от высших центров в спинной и продолговатый мозг. Эти же проводящие пути образуют ножки мозжечка и пучки волокон между скоплениями нервных клеток.
Черная субстанция отвечает за координацию актов глотания и жевания, а также участвует в регуляции пластического тонуса; имеет значение при выполнении пальцами мелких движений, требующих большой точности. Поэтому черная субстанция лучше развита у приматов.
Красное ядро — крупное скопление серого вещества среднего мозга, состоит из крупных и мелких клеток. Оно связано проводя-
 щими путями с мозжечком и ядрами полосатого тела, а также со §пинным мозгом (руброспинальный путь). Красные ядра с окружающей их ретикулярной формацией, несомненно, участвуют в регуляции мышечного тонуса. Можно сравнить поведение деце-|ебрированного животного, у которого со спинным и продолговатым мозгом сохранилась связь лишь посредством варолиева моста, мозжечка и нижнего участка среднего мозга без области красных ядер, с поведением так называемого таламического животного, у которого удалены кора мозга и подкорковые узлы, но средний мозг с его красными ядрами и зрительные бугры сохранены и остаются в связи с продолговатым мозгом.
щими путями с мозжечком и ядрами полосатого тела, а также со §пинным мозгом (руброспинальный путь). Красные ядра с окружающей их ретикулярной формацией, несомненно, участвуют в регуляции мышечного тонуса. Можно сравнить поведение деце-|ебрированного животного, у которого со спинным и продолговатым мозгом сохранилась связь лишь посредством варолиева моста, мозжечка и нижнего участка среднего мозга без области красных ядер, с поведением так называемого таламического животного, у которого удалены кора мозга и подкорковые узлы, но средний мозг с его красными ядрами и зрительные бугры сохранены и остаются в связи с продолговатым мозгом.
Тонические рефлексы ствола мозга.Важнейшая функция ствола мозга, в частности продолговатого и среднего мозга, состоит в перераспределении мышечного тонуса в зависимости от положения тела в пространстве. Перераспределение тонуса осуществляется рефлекторным путем и обеспечивает сохранение равновесия тела. Всю совокупность тонических рефлексов голландский ученый Р. Магнус разделил на две большие группы: статические рефлексы; статокинетические рефлексы.
Статические рефлексы.Выражены при спокойном стоянии, лежании и сидении в различных позах, они обусловливают определенное положение тела в пространстве. Такие рефлексы делятся на две большие группы. Первая группа обеспечивает определенное положение или позу тела — это рефлексы положения, или по-зотонические рефлексы. Вторая же способствует возвращению тела из неестественного положения в нормальное — это установочные, или выпрямительные, рефлексы.
Позотонические рефлексы включают в себя лабиринтные и шейные. Лабиринтные рефлексы, для осуществления которых достаточно наличия продолговатого и спинного мозга, обеспечивают фиксирование конечностей в определенном положении. В эксперименте на шею децеребрированного животного надевали гипсовую повязку, чтобы голова сохраняла постоянное положение по отношению к туловищу. При этом не происходит раздражение проприорецепторов шеи. Затем животное вращали вокруг оси, условно проведенной через оба ^иска. В положении, когда животное было спиной вниз, тонус разгибателей становился максимальным и конечности вытягивались, а если же оно оказывалось спиной вверх, то тонус этих мышц снижался и конечности сгибались.
На тонус всех мышц помимо лабиринтных влияют также шейные рефлексы. Влияние шейных рефлексов сказывается в том, что при поворотах головы по отношению к туловищу справа налево или слева направо тонус разгибателей максимален на той стороне тела, к которой обращено темя. Если же поворачивать голову вокруг оси, условно проведенной через оба слуховых прохода, то при наклоне головы вниз тонус разгибателей увеличивается в зад-
них и снижается в передних конечностях. При запрокидывании головы назад, наоборот, тонус разгибателей передних конечностей увеличивается, а задние конечности сгибаются.
Рефлексы с лабиринтов на шейные мышцы. Шейные и лабиринтные тонические рефлексы осуществляются в связи друг с другом, так как импульсы с лабиринтов влияют на тонус шейных мышц. Максимум тонуса шейных мышц наблюдается при тех положениях головы, когда импульсы с лабиринтов ведут к рефлекторному усилению тонуса разгибателей конечностей. Импульсы с каждого лабиринта влияют преимущественно на шейные мышцы противоположной стороны (поэтому при разрушении лабиринта голова животного повернута в сторону повреждения).
Выпрямительные рефлексы осуществляются средним мозгом и, следовательно, отсутствуют у бульбарных животных. В осуществлении этих рефлексов участвуют рецепторы лабиринтов, шейных мышц и кожи.
Если мезенцефальное животное положить на бок, оно поднимает голову и устанавливает ее в естественное положение теменем кверху. Это происходит в результате возбуждения рецепторов вестибулярного аппарата. Но даже при разрушении вестибулярного аппарата голова выпрямляется в том случае, если тело лежит на боку на какой-нибудь твердой поверхности. В этом случае в ответ на одностороннее раздражение кожи происходит рефлекторное выпрямление головы. Если на это же животное положить сверху доску с небольшим грузом, то происходит симметричное двустороннее раздражение кожных рецепторов и голова снова падает вниз. Если доску убрать, то голова рефлекторно снова поднимается.
Подъем головы —это только первая фаза выпрямительных рефлексов. Вторая фаза заключается в рефлекторном выпрямлении туловища, которое следует за подъемом головы. Этот рефлекс возникает от раздражения проприорецепторов мышц шеи и рецепторов кожи туловища.
Статокинетические рефлексы.Начинаются с одного рецептивного поля — с рецепторов полукружных каналов внутреннего уха. Возникают при угловом или линейном ускорении.
При вращении животного наблюдается нистагм головы, т. е. медленный поворот в сторону, противоположную вращению, и быстрый возврат в исходное положение. Глаза реагируют на вращение аналогичным образом — глазной нистагм. Осуществление статокинетичес-ких рефлексов возможно только при участии ядер среднего мозга.
РЕТИКУЛЯРНАЯ ФОРМАЦИЯ
В центральной части ствола мозга находится образование, состоящее из диффузных скоплений клеток различных типов и размеров, которые густо переплетаются множеством волокон, идущих в разные направления. Внешний вид нервной ткани этой об-
37 — 3389
ласти под микроскопом напоминает сеть. Дейтерс, впервые описавший ее строение в конце XIXв., назвал ее сетчатой или ретикулярной формацией. Такого разнообразия ядер нет ни в одном отделе ЦНС, что говорит о многообразии ее функций. В ретикулярной формации ствола мозга человека, кошки и кролика выделено множество отдельных ядер и клеточных групп.
В медиальной части продолговатого мозга лежит ретикулярное гигантоклеточное ядро, которое занимает около двух третей ретикулярной формации. Спереди от него располагается ядро лицевого нерва, а сзади — ретикулярное каудальное ядро, граничащее со спинным мозгом. Гигантоклеточное ядро отличается полиморф-ностью своих составных частей. Кроме гигантских в нем имеются крупные, средние и мелкие нейроны. В других ретикулярных ядрах гигантские нейроны отсутствуют.
Продолжением гигантоклеточного ядра в варолиевом мосту является каудальное ретикулярное ядро моста. Спереди это ядро переходит в оральное ретикулярное ядро моста, которое, в свою очередь, — в клеточную группу, образующую ретикулярную формацию среднего мозга.
В ретикулярную формацию поступает множество афферентных волокон, при этом три ретикулярных ядра, дающие проекции в мозжечок, получают большое количество афферентных волокон из его ядер. Спинно-ретикулярные волокна начинаются из нейронов задних и передних рогов спинного мозга и столбов Кларка, а оканчиваются в медиальных ядрах ретикулярной формации продолговатого мозга и варолиева моста. Кортикоретикулярные волокна возникают из различных областей коры, но преимущественно из лобной доли.
По своим эфферентным связям ядра ретикулярной формации делятся на две группы. Одна из этих групп, состоящая из ретикулярного ядра покрышки моста, латерального и парамедиальных ретикулярных ядер, проецируется в мозжечок. Ядра медиальных областей ретикулярной формации, составляющие вторую группу, посылают свои волокна к другим ядрам ретикулярной формации, к моторным и сенсорным ядрам черепно-мозговых нервов, к нейронам среднего и промежуточного мозга, к различным нейронам коры и спинного мозга.
Ретикулярная формация состоит из двух частей: нисходящей и восходящей. По нисходящим путям из среднего и продолговатого мозга к мотонейронам спинного мозга передаются импульсы либо активирующие, либо тормозящие функцию скелетных мышц. Стимуляция медиальных областей ретикулярной формации продолговатого мозга приводит к торможению моно-синаптических и полисинаптических рефлексов спинного мозга у анестезированных кошек. Основной поток нисходящих влияний на сегментарную деятельность спинного мозга посылает ретикулярное гигантоклеточное ядро.
Стимуляция латеральных областей ретикулярной формации варолиева моста и среднего мозга вызывает мощное генерализованное усиление моторной активности спинного мозга. Усиление или торможение моторной активности может происходить при раздражении одних и тех же участков мозга, причем тот или иной эффект зависит от интенсивности и продолжительности раздражения ствола мозга.
Ретикулярная формация оказывает модулирующее влияние на возбудимость афферентных переключений, а также передачу афферентных импульсов в сенсорных ядрах ствола головного мозга. Гигантоклеточные и мелкоклеточные ядра рассматриваются как центр вдоха. Медиальная ретикулярная формация не содержит нейронов, принадлежащих собственно дыхательному центру, однако границы, указываемые для расположения респираторных нейронов, захватывают области гигантоклеточного и мелкоклеточного ядер ретикулярной формации, а также область ретикулярного латерального ядра. Инспираторные и экспираторные нейроны, локализованные в соответствующих зонах продолговатого мозга, посылают свои аксоны в спинной мозг. Большинство аксонов респираторных нейронов перекрещивается в нижних частях продолговатого мозга и вступает в спинной мозг вместе с частью не-перекрещивающихся волокон.
Ретикулярная система влияет на деятельность сердца, сосудов, органов пищеварения, выделения и т. д. Большое значение имеет ее воздействие на железы внутренней секреции, особенно гипофиз.
Ретикулярная формация оказывает мощное влияние ив восходящем направлении — на кору больших полушарий головного мозга. Стимуляция каудальных и ростральных отделов ретикулярной формации ствола мозга спящего животного приводит к обширной активации коры, выражающейся в десинхрониза-ции электроэнцефалограммы (ЭЭГ), и пробуждению спящего животного, тогда как отключение восходящих ретикулярных путей от коры вызывает у животных глубокий сон. Полученная во время глубокого сна ЭЭГ сходна с таковой при коме или ступоре. Де-синхронизация ЭЭГ — реакция активации — происходит в результате возбуждения ретикулярной формации под воздействием любой афферентной системы.
При поражениях только сенсорных трактов явление комы не развивается. Это подтверждает тот факт, что восходящие конические влияния на кору мозга исходят из ретикулярной формации ствола мозга, а именно варолиева моста.
Деятельность восходящих путей поддерживается импульсами от всех рецепторов организма, так как от афферентных волокон, несущих импульсы к таламусу, отходят многочисленные коллатерали к ретикулярной формации. Кроме того, ретикулярная формация получает импульсы от мозжечка, подкорковых ганглиев, гипоталамуса, лимбической системы и коры больших полушарий.
37*
| Молекулярный слой |
| Зернистый слой |
 С сетчатой системой связано проявление различных эмоций. Разрушение очень небольшого ее участка в гипоталамусе превращает послушное, прирученное животное в дикое. Раздражение же соответствующего участка ретикулярной формации может вызвать чувство страха наказания или удовольствие.
С сетчатой системой связано проявление различных эмоций. Разрушение очень небольшого ее участка в гипоталамусе превращает послушное, прирученное животное в дикое. Раздражение же соответствующего участка ретикулярной формации может вызвать чувство страха наказания или удовольствие.
Оказывая большое влияние на кору и поддерживая уровень активности, ретикулярная формация вместе с тем находится под постоянным регулирующем влиянием коры головного мозга.
МОЗЖЕЧОК
Мозжечок представляет собой образование, расположенное позади больших полушарий мозга над продолговатым мозгом и варо-лиевым мостом. В эволюционном плане это очень древняя структура. Он участвует в координации всех сложных двигательных актов организма, включая и произвольные движения.
Мозжечок включает в себя среднюю часть — червячок, по бокам которого расположены два полушария и две небольшие боковые доли. Верхняя поверхность полушарий мозжечка представляет собой серое вещество — это кора мозжечка. В сером веществе различают три слоя: поверхностный, или молекулярный; слой клеток Пуркинье и внутренний — гранулярный. В белом веществе мозжечка находятся отдельные скопления серого вещества — ядра мозжечка. В каждом полушарии имеется по три ядра: зубчатое, пробковидное и шаровидное.
Связь мозжечка с другими отделами ЦНС осуществляется посредством большого количества нервных волокон, образующих толстые пучки: нижние, средние и верхние ножки мозжечка (рис. 13.15). Через нижние ножки проходят пучки Флексига и волокна находящихся в продолговатом мозге ядер пучков Голля и Бурдаха, а также пучки от вестибулярного ядра. Через средние ножки к мозжечку поступают волокна, несущие импульсы от коры больших полушарий; через верхние ножки вступают пучки Говер-са и волокна, идущие от переднего четверохолмия.
Эфферентные волокна, идущие от мозжечка, начинаются в его центральных ядрах. Они проходят через верхние ножки мозжечка и заканчиваются с перекрестом в красном ядре среднего мозга, в ядрах таламуса и гипоталамуса, в ретикулярной формации ствола мозга, в подкорковых ядрах и в продолговатом мозге.
Таким образом, мозжечок связан афферентными и эфферентными волокнами со всеми отделами ЦНС.
Наиболее широко используемым подходом для анализа функций мозжечка явился метод его удаления (экстирпация). Первые такие опыты были проведены еще в конце XIX в. Лючиани. Он установил после экстирпации три постепенно переходящие друг в друга стадии нарушения движения:
\мв v
Рис. 13.15. Схема синаптических связей нейронов мозжечка (Дж. Эккле, 1969)
Показаны возбуждающие синапсы, образуемые на грушевидных нейронах (клетки Пуркинье) лазающими волокнами (ЛВ), аксонами клеток-зерен (КЗ), которые, в свою очередь, активируются мшистыми волокнами (MB), и тормозные синапсы, образованные аксонами звездчатых (ЗвК) и корзинчатых клеток (Т— торможение; В— возбуждение)
стадия раздражения. Обусловлена не только удалением мозжечка, но и травмированием прилежащих к нему отделов мозга, внутричерепным кровоизлиянием, отеком тканей;
стадия выпадения функций. Выявляются характерные симптомы удаления мозжечка;
стадия компенсации. Несколько выравниваются нарушенные двигательные реакции.
В первые дни после операции полного удаления мозжечка животное совершенно беспомощно. Постепенно его движения восстанавливаются, но остаются беспорядочными, оно шатается, падает. В положении стоя для сохранения устойчивости животное широко расставляет конечности.
После удаления одной половины мозжечка конечности соответствующей стороны сильно вытянуты, животное двигается по кругу — это так называемые манежные движения.
Таким образом, удаление или поражение мозжечка вызывает расстройство статических и статокинетических рефлексов. Это указывает на то, что влияние мозжечка распространяется на тонические рефлексы положения тела и установочные рефлексы, осуществляемые центрами ствола мозга, а также на моторную зону коры больших полушарий и связанные с ней нервные центры.
При этом наблюдаются нарушения функций мозжечка.
Атония — резкое понижение тонуса мышц, которое возникает после удаления мозжечка. Через какой-то промежуток времени тонус некоторых мышц может повышаться. Всестороннее изучение функций мозжечка проводил Л. А. Орбели. Очень часто удаление мозжечка проявляется не в виде атонии, а нарушением регуляции мышечного тонуса — дистонии.
Атаксия — нарушение координации движений: их точности, скорости и направления (дисметрии). Атаксия проявляется резко выраженным расстройством походки. Конечности при движении поднимаются очень высоко и с силой ударяются о пол. Прежде чем попасть мордой в кормушку, животное несколько раз наклоняет и запрокидывает голову, как маятник.
Астазия — неспособность мышц к титаническим сокращениям. В результате голова, туловище и конечности животного дрожат. Особенно отчетливо эти явления проявляются после движения животного.
Астения — утомление вследствие повышения обмена веществ при очень быстрой работе мышц, которое связано с тем, что движения производятся несоразмерно и при участии большого количества мышц.
Астезия — потеря чувствительности.
Дезэквилибрация — нарушение равновесия. Явно проявляется у обезьян при удалении определенной доли мозжечка, тесно связанной с ядрами продолговатого мозга. После такой операции не происходит нарушения рефлексов положения тела в пространстве и произвольных движений. Пока такое животное лежит, у него не видно никаких нарушений, так как оно способно самостоятельно брать пищу руками и подносить ее ко рту, но сидеть оно может, только прислонясь спиной к опоре, а стоять самостоятельно не способно. Через длительное время после удаления мозжечка все эти симптомы несколько ослабевают, но не исчезают полностью.
Кроме регуляции двигательных функций мозжечок оказывает влияние на вегетативные функции. Раздражение его часто сопровождается симпатическими эффектами: расширением зрачков, повышением артериального давления, учащением пульса, восстановлением работоспособности утомленных мышц. После экстирпации мозжечка ослабляется моторная функция кишечника, тормозится секреция желудочного и кишечного сока.
ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг состоит из таламуса, или зрительных бугров, гипоталамуса и эпиталамуса. Промежуточный мозг образует стенки третьего желудочка. В процессе эмбриогенеза он формируется вместе с большими полушариями из переднего мозгового пузыря.
Таламус, или зрительные бугры, — парное образование. Через него к коре поступает вся информация от рецепторов, воспринимающих раздражение из внешней и внутренней среды организма. В ядрах таламуса происходит тесное взаимодействие импульсов и их подкорковая интеграция. В таламусе различают три области — переднюю, латеральную и медиальную. Все ядра (около 40) таламуса подразделяют на специфические и неспецифические.
К специфическим ядрам таламуса принадлежат переключающие и ассоциативные ядра. Группа латеральных талами-ческих ядер служит передаточным образованием для тактильных и кинестетических импульсов, поступающих по волокнам медиальной петли. К переключающим относятся передние (дорсальное, вентральное и медиальное), вентролатеральное, заднее вентральное ядра и коленчатые тела.
Латеральное коленчатое тело является переключающим ядром зрительных сигналов. К нейронам этого ядра поступают импульсы от первичных зрительных центров переднего четверохолмия. Отростки нейронов латерального коленчатого тела направляются в зрительную зону коры больших полушарий.
Медиальное коленчатое тело является переключающим ядром слухового пути. Здесь расположены нейроны, к которым поступают импульсы от первичных слуховых центров заднего четверохол- * мия. Отростки нейронов медиального коленчатого тела идут в слуховую зону коры больших полушарий.
Импульсы, поступающие к таламусу по волокнам от ядер Голля и Бурдаха в продолговатом мозге и по спинно-таламическому пути, а также по волокнам от ядер тройничного нерва, доставляют информацию от рецепторов кожи лицевой части головы, туловища, конечностей и от проприорецепторов. Эта информация поступает в заднее вентральное ядро таламуса и далее по его нейронам в заднюю центральную извилину коры больших полушарий — в соматосенсорную зону.
Передние ядра таламуса получают импульсы от висцерорецеп-торов и частично от обонятельных рецепторов, далее импульсы идут в лимбическую область больших полушарий.
Ассоциативные ядра таламуса, расположенные в его передней части, получают импульсы от переключающих ядер и передают их в ассоциативные зоны мозговой коры. К ассоциативным ядрам относятся латеральные, медиодорсальное и пушечное ядра. Латеральные ядра таламуса передают импульсы в теменную область коры больших полушарий.
Неспецифические ядра таламуса по своему происхождению более древние; они включают в себя срединные и инт-раламинарные ядра, а также медиальную часть переднего вентрального ядра. Нейроны неспецифических ядер сначала передают сигналы в подкорковые структуры, от которых импульсация поступает одновременно в разные отделы коры. Неспецифические
ядра служат продолжением ретикулярной формации среднего мозга, представляя собой ретикулярную формацию таламуса.
Таламус отвечает за возникновение ощущений: в его ядрах происходит переработка получаемой информации, в результате которой изменяется характер ощущений. При некоторых поражениях таламуса у животных появляются мучительные болевые ощущения, так, незначительное раздражение, прикосновение к коже, слабый укол вызывают приступы сильнейшей боли. До недавнего времени таламус считался высшим центром болевой чувствительности, но согласно современным данным высший центр болевых ощущений расположен в коре больших полушарий, а таламический болевой центр является промежуточным. Зрительные бугры участвуют и в регуляции мышечного тонуса: при перегрузке мозга по передней границе таламуса мышечный тонус повышается.
Гипоталамус,или подбугорье, образован группой небольших ядер, расположенных у основания мозга вблизи гипофиза под зрительными буграми. Клеточные ядра, образующие гипоталамус, представляют собой высшие подкорковые центры вегетативной нервной системы.
Нейронные образования гипоталамуса формируют преоптичес-кую, переднюю, среднюю, наружную и заднюю группы ядер. В пре-оптическую группу входят перивентрикулярное, медиальное и латеральное преоптические ядра. Переднюю группу ядер гипоталамуса образуют супрооптическое, супрохиазматическое и пара-вентрикулярное ядра, среднюю — вентромедиальное и дорсомеди-альное ядра. К наружной группе ядер относятся латеральное гипо-таламическое ядро и ядро серого бугра. Наконец, в заднем гипоталамусе различают заднее гипоталамическое, перефорникальное, премамиллярное, медиальное мамиллярное, супромамиллярное и латеральное мамиллярное ядра. Организация гипоталамуса характеризуется обширными и очень сложными афферентными и эфферентными связями.
Гипоталамус является диэнцефальным концом ретикулярной формации, что определяет их сходство: общий источник происхождения; наибольшая концентрация адреналина и ацетилхолина; на нейронах конвергируют афферентные импульсы. Однако эти структуры резко отличаются друг от друга по соотношению нейро-секреторной активности.
Гипоталамус участвует в терморегуляции: раздражение задних ядер приводит к гипертермии. В области средних и боковых ядер нейроны образуют центры насыщения и голода; стимулом служат отклонения в химическом составе притекающей крови. При недостатке в организме воды вследствие активации гипо-таламических зон появляется чувство чрезмерной жажды — полидипсия.
В гипоталамусе расположены центры, связанные с регуляцией
полового созревания. В опытах с вживлением электродов в задний гипоталамус животные вызывают самораздражение с высокой частотой в течение длительного времени. Гипоталамус также принимает участие в процессе чередования сна и бодрствования.
Гипоталамическая область находится в тесной морфологической и функциональной связи с гипофизом. Особенно это проявляется в регуляции водно-солевого обмена. Установлено, что перерезка нервного пути, связывающего гипоталамическую область с задней долей гипофиза, ведет к несахарному мочеизнурению — полиурии. Почечные условные и безусловные рефлексы в норме, по-видимому, всегда осуществляются при участии надоптического ядра подбугровой области, эфферентные нейроны которого участвуют в иннервации гипофиза, регулируя отделение антидиуретического гормона. Поступление этого гормона в кровь уменьшается при поступлении воды в организм и увеличивается при водном голодании.
Гипоталамус тесно взаимодействует с задней долей гипофиза, который как бы подвешен к подбугорью на тонкой ножке. Между ними имеются обширные нервные и сосудистые связи. Задняя доля гипофиза, являющаяся по происхождению производным нервной системы, выделяет антидиуретический гормон и гормоны, регулирующие деятельность матки и функцию половых желез. Эти вещества продуцируются клетками супро-оптического ядра и передаются в кровь по их аксонам в результате генерации потенциала действия в нейрогипофиз. С передней долей гипофиза (аденогипофиз) и с его средней долей связь гипоталамуса осуществляется через кровь — нейрогумораль-ным путем.
Эпиталамуспредставляет собой обонятельный центр и служит местом расположения железы внутренней секреции — эпифиза.
ПОДКОРКОВЫЕ ЯДРА
Подкорковые, или базальные, ядра входят в состав переднего мозга и расположены внутри больших полушарий между лобными долями и промежуточным мозгом. Они являются структурами ядерного типа. К ним относятся хвостатое ядро и скорлупа, названные «полосатым телом» в связи с тем, что они соединены между собой небольшими перемычками серого вещества и разделены пучками нервных волокон. Вместе с бледным шаром вышеперечисленные образования формируют стриопаллидарную систему подкорковых ядер. Базальные ганглии имеют обширные связи с другими отделами головного мозга. К полосатому телу идут афферентные волокна от разных областей коры больших полушарий, мозжечка и таламуса.
Кортикальные волокна берут начало практически из всей новой коры и распределяются по всем зонам полосатого тела. Наибольшее число волокон берет начало от клеток сенсомотор-ной зоны коры, наименьшее — от клеток зрительной зоны коры. Окончания кортикальных волокон образуют синапсы преимущественно на дендритных отростках полосатого тела. Кортикальные волокна возбуждают нейроны полосатого тела, а медиатором в этих синапсах является глутамат.
Проекции из таламуса берут начало в интраламинарных тала-мических ядрах, которые устанавливают синаптические контакты с шипиками или основаниями дендритов нейронов скорлупы и хвостатого тела. Это возбуждающие синапсы. Волокна, поступающие из черной субстанции, оказывают тормозящее влияние на активность нейронов.
Ядра стриопаллидарной системы принимают активное участие в координации двигательной активности. Электрическое раздражение хвостатого ядра вызывает стереотипное движение головы и дрожание передних конечностей. Перерезка путей, идущих из черной субстанции в полосатое тело, вызывает у животных неподвижность, отказ от корма и питья, а также отсутствие ответных реакций на внешние раздражения.
У человека полосатое ядро участвует в процессах запоминания: его раздражение может вызвать нарушение обучения и памяти. Стриопаллитарная система способна оказывать тормозящее влияние на различные проявления двигательной активности, а также на эмоциональные компоненты двигательного поведения.
|
Просмотров 4019 |
|
|
