Главная Обратная связь
Дисциплины:
Архитектура (936)
Биология (6393)
География (744)
История (25)
Компьютеры (1497)
Кулинария (2184)
Культура (3938)
Литература (5778)
Математика (5918)
Медицина (9278)
Механика (2776)
Образование (13883)
Политика (26404)
Правоведение (321)
Психология (56518)
Религия (1833)
Социология (23400)
Спорт (2350)
Строительство (17942)
Технология (5741)
Транспорт (14634)
Физика (1043)
Философия (440)
Финансы (17336)
Химия (4931)
Экология (6055)
Экономика (9200)
Электроника (7621)

Систолический и минутный объемы крови у животных
|
|
| Ввд животных | Систолический объем, мл | Минутный объем, л |
Лошадь 400...850 20...30 (в покое)
120... 160 (при нагрузке)
Крупный рогатый скот 600 30...40
Мелкий рогатый скот 55 4
Свинья 70 10
Собака 14 1,5
При физической нагрузке увеличиваются и систолический, и минутный объемы кровотока. Например, у лошади минутный объем при беге возрастает в 60...80 раз. У тренированных лошадей такой объем кровотока устанавливается за счет увеличения систолического выброса и некоторого учащения сердечных сокращений. У нетренированных лошадей или ослабленных предшествующим заболеванием сердце не способно увеличить значительно систолический объем, и возрастание минутного объема достигается в основном за счет учащения его работы. Это крайне невыгодный режим адаптации сердца к нагрузке, так как увеличение частоты сокращений ведет к укорочению сердечного цикла, особенно диастолы. Это приводит к недостаточному наполнению полостей сердца кровью, уменьшению силы сокращений и затрудняет восстановительные биохимические процессы в сердечной мышце.
Электрокардиография.Электрокардиография (ЭКГ) —это метод регистрации электрических потенциалов, возникающих при работе сердца. Запись биотоков сердца называется электрокардиограммой.
В норме возбуждение охватывает все отделы сердца последовательно, поэтому на его поверхности возникает разность потенциалов между возбужденными и невозбужденными участками, достигающая Ю0...120мВ (милливольт). Благодаря электропроводности тканей организма силовые линии тока достигают поверхности тела, где разность потенциалов между возбужденными и невозбужденными участками сердца всего 1...3 мВ, что можно измерить, приложив к соответствующим участкам кожи электроды.
Сердце находится в грудной клетке хотя и посередине, но верхушка его направлена влево. Из-за такого несимметричного расположения электрическая ось сердца сдвинута и проходит примерно от правого предсердия к левому желудочку, поэтому электроды накладывают на точки, несимметричные относительно сердца.
В ветеринарии применяют различные способы наложения электродов, или отведения. Стандартный способ отведения биопотенциалов — наложение электродов на конечности следующим образом:
I отведение: пясти левой и правой грудных конечностей; реги
стрируются потенциалы предсердий;
II отведение: пясть правой грудной и плюсна левой тазовой ко
нечностей; регистрируется возбуждение желудочков;
III отведение: пясть левой грудной и плюсна левой тазовой ко
нечностей; регистрируется возбуждение левого желудочка.
Применяют также грудные отведения биопотенциалов: один электрод располагают на конечности, а другие шесть — на различных участках грудной клетки.
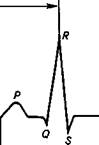
ЭКГ состоит из ровной изопотенциальной линии, которая соответствует потенциалу покоя, и пяти зубцов — Р, Q, R, S, Т. Гри зубца (Р, R и 7), идущие вверх от изопотенциальной ли-
17 — 3389
| циастолическшг |
| Интервал ORST •*-!—---------------- *> |

период Т-Р
Интервал QRS
>+-
Систолический период Р-Т
Рис. 6.6. Схема измерения интервалов ЭКГ:
Р— возбуждение предсердий; Q, R, S— возбуждение желудочков; R— начальная и 7"— конечная фазы возбуждения желудочков
нии, являются положительными, а два зубца {Q и S), направленные вниз от нее, — отрицательными (рис. 6.6).
Зубец Р— сумма потенциалов предсердий. Возникает в период распространения возбуждения по предсердиям. У крупных животных (лошадь, корова) зубец Р обьмно бывает раздвоенным, так как возбуждение вначале возникает в правом предсердии (в синусном узле) и только спустя 0,01 с достигает левого предсердия.
Интервал P—Q — время прохождения возбуждения от предсердий к желудочкам. Этот интервал соответствует атриовентрику-лярной задержке возбуждения.
Зубец Q — возбуждение внутренних слоев мышцы желудочков, правой сосочковой мышцы, перегородки, верхушки левого и основания правого желудочков.
Зубец R — распространение возбуждения на мышцы обоих желудочков.
Зубец S— охват возбуждением желудочков.
Интервал S—T отражает отсутствие разницы потенциалов в период, когда миокард охвачен возбуждением. В норме изопо-тенциален.
Зубец Т— фаза восстановления (реполяризации) миокарда желудочков.
QRS— время, в течение которого возбуждение успевает полностью охватить мышцы желудочков.
| Интервал R-R |
QRST— время возбуждения и восстановления миокарда желудочков.
Интервал Т—Р— возбуждение в желудочках уже закончилось, а в предсердиях еще не началось. Называется электрической диастолой сердца.
Интервал R—R (или Р—Р) соответствует полному циклу сердечной деятельности.
При анализе ЭКГучитывают высоту зубцов, их направленность от изопотенциальной линии и продолжительность интервалов.
ЭКГв комплексе с другими клиническими методами исследования применяется для диагностики заболеваний сердца, особенно таких, которые связаны с расстройством возбудимости и проводимости сердечной мышцы.
ЭКГиспользуется и в физиологических исследованиях для изучения состояния сердечно-сосудистой системы. Накоплена огромная база данных, раскрывающая видовые и индивидуальные особенности ЭКГу лошадей, крупного и мелкого рогатого скота, свиней, собак, верблюдов. Внедрение телерадиометрических методов регистрации биопотенциалов сердца позволило изучить ЭКГ у диких и полудиких животных, а также в экстремальных условиях — космические полеты, погружение в воду и др. Разработана также методика регистрации ЭКГу плода, что позволяет изучать развитие сердца, его функций и выявлять раннюю внутриутробную патологию.
РЕГУЛЯЦИЯ РАБОТЫ СЕРДЦА
Ритмические сокращения сердца обусловлены электрическими потенциалами, возникающими в пейсмекерах первого порядка, т. е. в синусном узле. Такая частота работы сердца называется синусной и наблюдается у животных в состоянии физиологического покоя. Сила сокращения сердца в определенном диапазоне, главным образом в состоянии покоя, также регулируется внутрисердечными механизмами (законы Боудича и Франка — Старлинга).
Однако при различных внешних воздействиях или при изменениях деятельности отдельных органов и систем организма работа сердца изменяется. В результате изменения, или регуляции, сердечной деятельности обеспечивается соответствие между уровнем обменных процессов в органах и количеством крови, нагнетаемой сердцем в сосудистую систему. Такое динамическое равновесие между потребностью организма и его реализацией достигается за счет изменения частоты и силы сокращений сердца, что связано уже не только с внутрисердечными механизмами, но и с внесер-дечными, или экстракардиальными.
Внутрисердечные механизмывключают в себя внутриклеточные механизмы, межклеточное взаимодействие и внутрисердечные периферические рефлексы.
17*
 Внутриклеточные механизмы направлены на усиление или ослабление синтеза клеточных белков, а также внутриклеточных структур, обеспечивающих биохимические процессы в клетках. Стимуляция этих процессов лежит в основе физиологической или рабочей гипертрофии сердечных мышц (например, у спортивных лошадей). К внутриклеточным механизмам применим закон Франка — Стерлинга (сила сокращения миокарда пропорциональна исходной длине его мышечных волокон).
Внутриклеточные механизмы направлены на усиление или ослабление синтеза клеточных белков, а также внутриклеточных структур, обеспечивающих биохимические процессы в клетках. Стимуляция этих процессов лежит в основе физиологической или рабочей гипертрофии сердечных мышц (например, у спортивных лошадей). К внутриклеточным механизмам применим закон Франка — Стерлинга (сила сокращения миокарда пропорциональна исходной длине его мышечных волокон).
Механизмы межклеточного взаимодействия обеспечиваются вставочными дисками, объединяющими клетки миокарда в функциональный синцитий. Благодаря вставочным дискам клетки обмениваются информацией и возбуждение очень быстро, практически одновременно охватывает всю сердечную мышцу.
Внутрисердечные периферические рефлексы — более высокий уровень регуляции. Эти рефлексы замыкаются не в центральной нервной системе, а во внутрисердечных нервных ганглиях, поэтому они имеют короткую рефлекторную дугу. Афферентные (чувствительные) нейроны воспринимают растяжения мышечных волокон сердца и коронарных сосудов, эфферентные (двигательные) нейроны иннервируют миокард и гладкие мышцы коронарных сосудов. Например, при переполнении кровью и растяжении правого предсердия, что обычно бывает при венозных застоях в большом круге кровообращения, усиливаются сокращения левого желудочка. Значение этого рефлекса в том, чтобы больше крови перекачать из полых вен в артерии. Однако так происходит только при нормальном или пониженном давлении крови в аорте. Если же камеры сердца переполнены кровью, а давление в аорте повышено, то растяжение предсердий на этом фоне угнетает сокращения миокарда и в аорту выбрасывается меньше крови. При нормальном притоке крови к сердцу, но высоком артериальном давлении сила сокращений миокарда увеличивается, способствуя «проталкиванию» крови в артерии.
Подобные внутрисердечные рефлексы стабилизируют наполнение кровью артериального русла: уменьшают выброс крови в артерии при переполнении их кровью и увеличивают при недостаточном возврате крови к сердцу.
Внесердечная,или экстракардиальная, регуляция сердечной деятельности—еще более высокий уровень приспособления сердца к потребностям организма. Она осуществляется нейрогуморальными механизмами, т. е. при участии ЦНС и биологически активных веществ, доставляемых к сердцу кровью.
Нервная регуляция. Нервная, или рефлекторная, регуляция осуществляется по принципу рефлексов. Это значит, что изменения работы сердца происходят вследствие действия каких-либо раздражителей по классической рефлекторной дуге: рецепторы — афферентные нервы — нервный центр — эфферентные нервы — сердце.
Рассмотрение нервной регуляции работы сердца целесообразно начать с эфферентной части рефлекторной дуги. Эфферентные, или центробежные, нервы сердца относятся к вегетативной нервной системе. Каждое вегетативное нервное волокно образовано двумя нервными клетками. Первый нейрон находится в центральной нервной системе и не доходит до эффектора, а его аксон заканчивается в нервном ганглии и называется преганглионарным. Второй нейрон — постганглионарный, начинается из нервного ганглия, а его аксон доходит до мышечных волокон сердца.
Парасимпатические преганглионарные нервные волокна начинаются в продолговатом мозге и идут в составе блуждающего нерва вагуса, являющегося десятой парой черепно-мозговых нервов (рис. 6.7.). Аксоны преганглионарных парасимпатических волокон заканчиваются в интрамуральных ганглиях сердца, главным образом в правом предсердии. Отсюда короткие аксоны пост-ганглионарных нейронов направляются к синусному и атриовентри-кулярному узлам проводящей системы сердца, и лишь небольшое количество волокон расходится в миокарде предсердий.
Симпатические преганглионарные нейроны находятся в шести первых сегментах грудной части спинного мозга, а их аксоны заканчиваются в шейных и грудных симпатических узлах. Из шейного звездчатого ганглия, имеющего важное значение в их распределении, выходят постганглионарные симпатические волокна, иннервирующие главным образом мышцы желудочков и в меньшей степени мышцы предсердий.

|
Вегетативные нервы влияют на все свойства сердечной мышцы. Влияние на частоту сокращений сердца называется хронотроп-ным, на силу сокращений — инотропным, на возбудимость — бат-мотропным, на скорость распространения возбуждения — дромо-тропным. Если работа сердца при воздействии на него урежается или ослабляется, говорят об отрицательном влиянии, а если учащается или усиливается — о положительном.
При раздражении симпатических нервов наблюдаются положительные хронотропный, ино-гропный, батмотропный и дромотропный эффекты. Раздражение парасимпатических нер-нов вызывают отрицательные воздействия, т. е. угнетение работы сердца вплоть до его остановки (рис. 6.8). Открытие тормозного действия вагуса на сердце лягушки впервые пока-
Рис. 6.7. Схема иннервации сердца:
/ — кора мозга; 2— продолговатый мозг; 3, 4— симпатические во-иокна (пост- и преганглионарные); J—шейные симпатические ушы;6— волокна блуждающего нерва (пре- и постганглионарные); 7— промежуточный мозг

|
зало, что нервная система может не только усиливать функцию органа, но и снижать.

|

|
| Рис. 6.8. Сокращения сердца при раздражении блуждающего (А) и симпатического (Б) нервов; а — отметка раздражения |
В составе как симпатических, так и парасимпатических нервов обнаружены путем тщательной препаровки и последующей стимуляции электрическим током отдельные волокна, которые оказывают или хронотропные, или инотропные влияния. Влияние нервов на сердце реализуется через нервные медиаторы. В синап-тических окончаниях пре- и пост-ганглионарных волокон парасимпатических нервов выделяется ацетилхолин, такие волокна называются холинергическими. Ацетилхолин выделяется также в окончаниях преганглионарных симпатических волокон — в симпатических ганглиях. В окончаниях постганглионарных симпатических волокон освобождается норадреналин, это — адренерги-ческие волокна.
Вызываемый при раздражении нервов эффект зависит не только от медиаторов, их химической структуры, но и от белковых молекул — рецепторов, находящихся на постсинаптических мембранах. Именно взаимодействие медиатора с рецептором определяет ответную реакцию тканей на раздражение. Один и тот же медиатор может вызывать противоположные реакции в различных тканях. Например, в скелетных мышцах ацетилхолин является возбуждающим медиатором, а в сердце — тормозным.
Механизм тормозного влияния вагуса на сердце связан с тем, что ацетилхолин, освобождающийся из пресинаптической мембраны, взаимодействуя с холинрецептором, вызывает повышение калиевой проницаемости мембран пейсмекерных клеток и их гиперполяризацию. Вследствие этого новый потенциал действия возникает позднее. Укороченный потенциал действия не способен возбудить достаточное количество кардиомиоцитов. Помимо этого гиперполяризация мышечных волокон противодействует входу кальция внутрь миофибрилл. Ацетилхолин также угнетает активность АТФ и снижает силу сокращения саркомеров. Таким образом, раздражение вагуса приводит к повышению порога раздражимости предсердий, подавлению автоматии и замедлению проводимости атриовентрикулярного узла и уменьшению силы сокращения миокарда (см. рис. 6.8, А).
Если раздражение вагуса продолжать длительное время, то остановившееся вначале сердце начинает вновь сокращаться, причем сокращения оказываются более сильными и редкими, чем в
норме. Такое явление называется «ускользанием сердца из-под влияния вагуса», и, возможно, оно является результатом компенсаторного усиления симпатических влияний на сердце. Есть и другая точка зрения: угнетение вагусом автоматии синусного узла способствует проявлению других очагов автоматии, в первую очередь атриовентрикулярного узла.
Раздражение симпатических нервов (см. рис. 6.8, Б) вызывает учащение сердечного ритма и увеличение силы сокращений сердца. Механизмы влияния симпатических нервов на сердце также объясняются взаимодействием медиатора с рецепторами постсинаптических мембран. Норадреналин, выделяющийся из окончаний адренергических симпатических нервов, взаимодействует с бета-рецепторами поверхностной мембраны. В результате увеличивается проницаемость мембраны для натрия и кальция и уменьшается для калия. Скорость возникновения потенциала действия возрастает, увеличивается возбудимость и проводимость сердечной мышцы. Симпатические нервы улучшают трофику (питание) сердечной мышцы и расширяют коронарные сосуды, улучшая ее кровоснабжение.
Норадреналин разрушается в синапсах значительно медленнее, чем ацетилхолин, поэтому адренергические влияния продолжительнее холинергических и сохраняются некоторое время после прекращения симпатической стимуляции.
Если у животного перерезать парасимпатические нервы, идущие к сердцу, или заблокировать передачу возбуждения с парасимпатических нервов на миокардиоциты (например, введением атропина), то частота сокращений сердца резко возрастает —в полтора-два раза.Если же перерезать симпатические нервы или ввести бета-адреноблокатор, то частота сокращения сердца уменьшится, но лишь на 15...20 %. Эти наблюдения, неоднократно подтвержденные на лабораторных животных и на людях, показали, что в состоянии покоя в большем тонусе находится блуждающий нерв. Он постоянно тормозит работу сердца в большей или в меньшей степени. У новорожденных животных тонус блуждающих нервов отсутствует, поэтому у них частота сокращений сердца намного больше, чем у взрослых.
Тонус блуждающих нервов поддерживается постоянными импульсами, поступающими в продолговатый мозг, в ядра вагуса, от различных рецепторов. Особенно важное значение имеют рецепторы дуги аорты и каротидного синуса, которые раздражаются при каждом систолическом выбросе крови из левого желудочка в аорту.
При мышечной работе, психоэмоциональных нагрузках увеличивается тонус симпатической нервной системы, и это вызывает положительные инотропные и хронотропные влияния на сердце, но при этом тонус парасимпатических нервов снижается.
Таким образом, несмотря на противоположное влияние на сердце, между симпатическими и парасимпатическими центрами сер-
дечной деятельности нет антагонизма. Эти центры находятся между собой в состоянии реципрокного торможения, когда возбуждение одного отдела автоматически ведет к торможению другого. Поэтому в состоянии физиологического покоя преобладает тонус парасимпатических нервов и сердце работает на 1/5... 1/6 ниже своих возможностей. Зато при различных нагрузках повышается тонус симпатического отдела и сердце увеличивает частоту и силу сокращений в несколько раз.
Помимо продолговатого мозга, где расположены центры блуждающего нерва, и спинного мозга с центрами симпатических нервов в регуляции работы сердца участвуют вышележащие отделы мозга — средний и промежуточный, а также лимбическая система и кора больших полушарий. При раздражении этих отделов возникают разнообразные рефлекторные реакции, приспосабливающие работу сердца к потребностям организма. Так, кора больших полушарий участвует в образовании условных рефлексов (например, предстартовой тахикардии у спортивных лошадей), а также в проявлении различных эмоций.
Большая часть сердечных рефлексов сочетается с сосудистыми рефлекторными реакциями и направлена на регуляцию кровообращения в целом. Рефлекторные изменения работы сердца могут возникнуть при раздражении самых разнообразных экстеро- и интерорецепторов.
Рефлексы, начинающиеся с сосудистых рефлексогенных зон, имеют очень большое значение в регуляции сердечной деятельности, сосудистого тонуса и других функций организма.
Синокаротидная зона, или зона каротидного синуса, — парная, находится в разветвлениях правой и левой сонных артерий на наружные и внутренние ветви. Здесь имеется «каротидное тельце», или каротидный синус, представляющий собой скопление большого числа нервных клеток и чувствительных нервных окончаний. Каротидный синус имеет собственную оболочку и сеть питающих его кровеносных сосудов.
Рецепторы синокаротидной зоны чувствительны к колебаниям артериального давления, его изменения воспринимаются баро-или прессорецепторами. При повышении артериального давления импульсы от рецепторов по синусному нерву (веточка языкогло-точного нерва) передаются в продолговатый мозг и переключаются на ядра блуждающего нерва. По вагусу ответная команда поступает к сердцу, вызывая отрицательные инотропные и хронотроп-ные эффекты. В результате артериальное давление снижается. Если же артериальное давление ниже какого-то уровня, свойственного данному животному, барорецепторы не получают нужной информации, что ведет к ослаблению влияния блуждающего нерва на сердце и усилению симпатической стимуляции. В синокаротидной зоне имеются также хеморецепторы, чувствительные к
концентрации кислорода, диоксида углерода, водородных ионов, никотина и других веществ.
Вторая жизненно важная сосудистая рефлексогенная зона находится в дуге аорты. Здесь также расположено большое скопление баро- и хеморецепторов, раздражение от которых передается по аортальному, или депрессорному, нерву в продолговатый мозг, откуда по вагусу импульсы идут к сердцу, уменьшая его работу.
Таким образом, обе указанные зоны — аортальная и синокаротидная — функционируют однонаправленно, как бы дублируя друг друга. Аортальная зона контролирует общее артериальное давление в организме, а синокаротидная зона регулирует кровоснабжение мозга.
Большую роль в регуляции сердечной деятельности играет рефлексогенная зона вблизи устья полых вен, а также в правом предсердии. Рефлекс с барорецепторов этой зоны носит название рефлекса Бейнбриджа. Он заключается в том, что при переполнении кровью и растяжении полых вен импульсы от рецепторов этой зоны направляются по сегментарным спинномозговым нервам в грудную часть спинного мозга, где находятся центры симпатических нервов сердца. Симпатические нервы усиливают работу сердца, и давление крови в полых венах снижается.
Рефлекторные изменения работы сердца возникают и при раздражении других рецепторных участков (зон) организма. Рефлекс Гольца проявляется в виде резкого замедления, вплоть до остановки сердца, при раздражении механорецепторов брюшной стенки и органов брюшной полости, а также при резком охлаждении кожи живота. Рефлекс Ашнера заключается также в замедлении сокращений сердца при несильном (неболевом) надавливании на глазное яблоко. У здоровых людей при этом частота сокращений сердца уменьшается примерно на 10 ударов в минуту. Рефлекс Ашнера используют иногда для определения тонуса.вегетативной нервной системы. Рефлекс Парина проявляется следующим образом: при увеличении давления в легочной артерии ритм сокращений сердца замедляется.
Замыкание рефлекторных дуг указанных рефлексов происходит на уровне продолговатого мозга, это типичные вагусные рефлексы с отрицательными инотропным и хронотропным эффектами.
Усиление работы сердца происходит при болевых раздражениях, сокращении скелетных мышц во время физической нагрузки. В этих случаях рефлекторные реакции осуществляются через симпатическую нервную систему.
Очень чутко реагирует сердце на изменение температуры крови. При повышении температуры частота сокращений сердца возрастает, при снижении — уменьшается. Влияние тепла и холода сказывается прежде всего на возбудимости и проводимости сердца, особенно клеток синусного узла. Изменения частоты и силы
сокращений сердца наблюдаются даже при локальном охлаждении или согревании синусного узла. Кроме того, от температуры притекающей крови зависит скорость биохимических реакций.
Не всегда отмечаются однонаправленные изменения силы и частоты сокращений сердца. При учащении сердечного ритма укорачивается диастолическая пауза и сила сокращения может уменьшиться. Напротив, урежение сердечного ритма может привести к более мощным сокращениям, так как в этом случае увеличивается диастолическое наполнение сердца кровью.
Гуморальная регуляция сердца. Биологически активные вещества влияют либо непосредственно на сердечные волокна, либо на эфферентные нервные окончания синапсов, либо рефлекторно — через центральную нервную систему.
Катехоламины (адреналин, норадреналин) — гормоны, выделяющиеся мозговым веществом надпочечников и островками хром-аффиновой ткани, разбросанными по всему организму. Катехоламины являются также медиаторами адренергических нервов. Они действуют на сердце аналогично симпатической нервной системе, реагируют с бета-рецепторами, вызывая положительные инотроп-ные и хронотропные воздействия.
Ацетилхолин образуется в холинергических нервных окончаниях. К сердцу он поступает не только через синапсы парасимпатических сердечных нервов, но через кровь и интерстициальную жидкость, действуя так же, как и парасимпатические нервы (отрицательные эффекты).
Положительное инотропное действие на сердце оказывают глюкагон — гормон поджелудочной железы, тироксин — гормон щитовидной железы и кортикостероиды — гормоны коры надпочечников. Действие указанных гормонов на сердце реализуется либо через циклический аденозинмонофосфат (цАМФ), находящийся в мембране клеток, либо через усиление тонуса симпатической нервной системы.
Большое значение в регуляции работы сердца имеют электролиты. Катионы кальция повышают возбудимость клеток миокарда, активируют фосфорилазу, участвуют в механическом сокращении миофибрилл. Кальций усиливает сердечные сокращения, но при избытке его в крови сердце останавливается в состоянии систолы, так как он в миофибриллах тормозит разобщение актино-вых и миозиновых нитей.
Катионы калия в повышенной концентрации снижают потенциал покоя миокардиоцитов и увеличивают калиевую проницаемость мембран. При небольшом превышении концентрации калий увеличивает возбудимость сердца, а при высокой — замедляет его работу и останавливает в фазе диастолы. Это явление используют в кардиохирургии для остановки сердца. Снижение калия в крови ниже физиологической нормы активизирует автоматию сердца, но при этом возможно нарушение сердеч-
ного ритма из-за стимуляции потенциальных пейсмекеров (например, атриовентрикулярного узла).
На работу сердца влияют уровень кислорода, диоксида углерода и водородных ионов в крови. Снижение содержания кислорода в крови, увеличение диоксида углерода или ацидоз угнетают сократительную активность миокарда.
|
Просмотров 2167 |
|
|
