Главная Обратная связь
Дисциплины:
Архитектура (936)
Биология (6393)
География (744)
История (25)
Компьютеры (1497)
Кулинария (2184)
Культура (3938)
Литература (5778)
Математика (5918)
Медицина (9278)
Механика (2776)
Образование (13883)
Политика (26404)
Правоведение (321)
Психология (56518)
Религия (1833)
Социология (23400)
Спорт (2350)
Строительство (17942)
Технология (5741)
Транспорт (14634)
Физика (1043)
Философия (440)
Финансы (17336)
Химия (4931)
Экология (6055)
Экономика (9200)
Электроника (7621)

Глава 9. БИОЛОГИЧЕСКИЕ МЕМБРАНЫ
|
|
Важнейшим структурным компонентом клетки являются биологические мембраны, которые представляют собой плоские высокоорганизованные структуры, состоящие преимущественно из белков и липидов. Они выполняют следующие основные функции:
· барьерную. Клеточная мембрана отделяет содержимое клетки от внешней среды, обеспечивая формирование внутренней среды клетки;
· компартментализации. Эта функция является частным проявлением барьерной. Мембраны разделяют внутриклеточное пространство на ряд отсеков – компартментов. На их основе возникают отдельные внутриклеточные органеллы (рис. 42) – ядро, митохондрии, эндоплазматичес-кий ретикуулум, комплекс Гольджи, пероксисомы, лизосомы.

Рисунок 42 – Строение эукариотической клетки
(Т.Т.Березов, Б.Ф.Коровкин, 1990)
Компартментализация, характерная для клеток эукариот, позволяет пространственно разделять метаболические пути, связанные с противоположно направленными превращениями одних и тех же веществ (синтез жирных кислот происходит в цитоплазме, а распад – в митохондриях и др.).
Несмотря на то, что мембраны выполняют барьерную функцию, они не представляют собой инертные оболочки. Для биологических мембран характерно явление избирательной проницаемости. Это свойство связано с присутствием в их структуре особых транспортных систем, каналов и пор. Наличие этих мембранных образований позволяет клетке осуществлять обмен веществ с внешней средой, обеспечивая введение в нее питательных веществ (субстратов окисления и пластических материалов) и выведение продуктов обмена.
Мембраны служат опорной структурой для целого ряда ферментов (мембраносвязанных ферментов), функционирование которых вне мембран невозможно.
Гидрофобный слой мембраны представляет собой своеобразный растворитель для гидрофобных молекул.
Мембранам присущ и ряд специализированных функций: они могут играть роль электрического изолятора на нервных проводниках (миелиновые мембраны), трансформатора энергии (внутренняя митохондриальная мембрана), участвовать в генерации электрического импульса (мембраны возбудимых клеток), обеспечивать процесс световосприятия (клетки сетчатки глаза) и др.
Биологические мембраны представляют собой чрезвычайно сложно устроенные гетерогенные структуры, различающиеся по химическому составу и функциям. Однако для них характерен и целый ряд общих свойств.
Впервые мембраны были обнаружены с помощью электронного микроскопа. На электронных микрофотографиях они представляют собой плоские структуры толщиной 60–100 мкм, имеющие характерное трехслойное строение. На периферии располагаются электронно-плотные слои, а в центре – прозрачная для электронов область (рис. 43).

Рисунок 43 – Изображение биологической мембраны на электронных
микрофотографиях (периферические темные слои – электронно-плотные,
центральный светлый слой прозрачен для электронов)
Изучение химического состава мембран стало возможно лишь с разработкой методов фракционирования мембранных структур клетки. Существенный прогресс в этом вопросе наступил после изобретения ультрацентрифуги и внедрения ее в практику биохимических исследований. Современные методы выделения мембран основаны на их разделении по плотности в гравитационном поле, которое создается в роторе ультрацентрифуги.
Исследования очищенных клеточных мембран показали, что практик-чески все они состоят из двух основных компонентов – липидов и белков. Белки формируют периферические электронно-плотные области мембран, абсорбирующие на себе электронно-плотный материал, используемый при фиксации мембраны (окись осмия). Между этими слоями располагается слой, образованный липидами, который прозрачен для электронов.
Основу гидрофобного слоя мембраны составляют фосфолипиды. Как видно из представленной ниже схемы строения фосфатидилэтаноламина (рис. 44), его молекула образована гидрофильной головкой, состоящей из остатка глицерина, фосфорной кислоты и аминоспирта, к которой присоединены два гидрофобных жирнокислотных радикала.

Рисунок 44 – Молекула фосфатидилэтаноламина (а)
и ее схематическое изображение (б)
При смешивании фосфолипидов с водой образуется устойчивая суспензия. В ее основе лежат мицеллы. Мицеллы представляют собой определенным образом устроенные устойчивые в растворах фосфолипидные частицы, которые стабилизируются за счет многочисленных нековалентных связей (водородных, гидрофобных, Ван-дер-Ваальсовых). В зависимость от характера растворителя мицеллы могут иметь противоположную ориентацию фосфолипидных молекул (рис. 45).
Как видно из представленного рисунка, прямая мицелла устроена таким образом, что полярные группы фосфолипидов находятся в непосредственном контакте с молекулами воды, а неполярные (гидрофобные) жирнокислотные остатки удалены от контакта с ней. За счет этого возникает устойчивая система, обеспечивающая поддержание молекул фосфолипидов в воде и других полярных растворителях.


а б
Рисунок 45 – Строение прямых
(а – в воде) и обратных (б – в бензоле) мицелл
Помимо способности к мицеллообразованию, фосфолипиды обладают свойством образовывать в полярных растворителях плоские бислойные структуры. За счет этого они широко используются для получения искусственных фосфолипидных мембран.
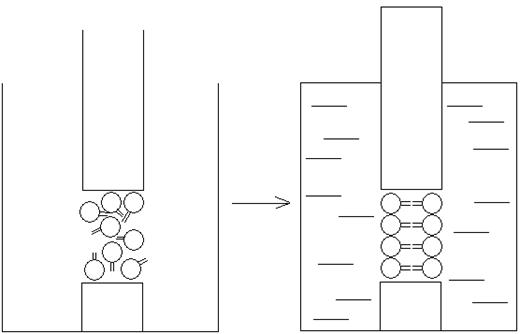
Для получения искусственных мембран в сосуде с полярным растворителем (водой) устанавливается перегородка из тефлона, в которой делается небольшое отверстие, диаметром около 1 мм. Перед заполнением сосуда водой в отверстие помещают каплю раствора фосфолипида в органическом растворителе. После заполнения сосуда происходит направленное перемещение молекул фосфолипида относительно молекул растворителя. В результате этого в отверстии тефлоновой пластинки возникает плоская структура, у которой в контакте с водой оказываются полярные головки молекул фосфолипидов, а их гидрофобные жирнокислотные хвосты оказываются изолированными от нее. Таким образом формируется искусственная фосфолипидная мембрана (рис. 46):
Искусственные мембраны обладают некоторыми свойствами, характерными для биологических мембран. Им присуща барьерная роль, они обладают избирательной проницаемостью, электрической емкостью и т.д. По этой причине они широко используются в экспериментальных исследованиях в качестве примитивной модели биологических мембран.
а б
Рисунок 46 – Получение искусственных фосфолипидных мембран
(расположение молекул фосфолипида в отверстии сухой тефлоновой
пластинки (а) и пластинки, помещенной в воду (б))
Строение и функции биологических мембран значительно отличаются от искусственных. Они более сложны и многообразны. Это связано со сложностью их строения. Если в состав искусственных фосфолипидных мембран входят исключительно фосфолипиды, то биологические мембраны содержат еще и другие представители липидов, а также белки и углеводы. В табл. 2 представлены сведения о химическом составе некоторых биологических мембран (миелина, эритроцитов и митохондрий).
Основу структуры биологической мембраны составляет липидный би-слой. Однако, как видно из табл. 2, в состав мембран, помимо липидов, входят и другие компоненты – белки и углеводы. В значительном количес-тве в структуру всех биологических мембран входит вода. По некоторым данным, ее содержание может достигать 20 % от массы мембраны.
Таблица 2 – Химический состав некоторых биологических мембран (%)
| Объект | Белки | Липиды | Углеводы |
| Милин | |||
| Эритроцитарная мембрана | |||
| Наружная митохондриальная мембрана | |||
| Внутренняя митохондриальная мембрана | - |
Липидные компоненты биологической мембраны представлены преимущественно фосфолипидами, которые формируют ее липидный бислой. Основными представителями мембранных фосфолипидов являются фосфатидилхолин и фосфатидилэтаноламин. В сумме их содержание может сос-тавлять до 80 % от содержания всех мембранных фосфолипидов. В значительно меньшем количестве в состав мембран входят фосфатидилсерин, фосфатидилинозитол, фосфатидная кислота, кардиолипин и сфингомиелин.
В небольшом количестве в состав липидного бислоя биологических мембран входят лизофосфатиды, которые представляют собой продукты частичного гидролиза фосфолипидов. В отличие от фосфолипидов в их сос-тав входит остаток только одной высшей жирной кислоты. Особое распространение среди лизофосфатидов в мембранах имеют лизофосфатидилхолин и лизофосфатидилэтаноламин.

Лизофосфатидилэтаноламин
При некоторых состояниях содержание лизофосфатидов в мембранах существенно возрастает. Это приводит к существенному изменению физи-ческих свойств липидного бислоя, делая его более проницаемым.
Для мембранных фосфолипидов характерен ряд общих свойств, которые касаются включенных в их состав жирнокислотных остатков. Их жирнокислотный состав отличается большой вариабельностью. Однако все жирные кислоты, включенные в состав мембранных фосфолипидов, имеют четное число углеродных атомов (от 12 до 26). В их радикале могут быть и двойные связи, количество которых в отдельных случаях достигает шести.
В состав фосфолипидов могут одновременно входить как насыщенные, так и ненасыщенные жирнокислотные остатки. При этом жирная кис-лота, содержащая в своей структуре двойную связь, как правило, связана сложной эфирной связью со вторым углеродным атомом глицерина. Наиболее часто ненасыщенные жирные кислоты входят в состав фосфатидилэтаноламина, фосфатидилсерина и фосфатидилинозитола. В то же время фосфатидилхолин и сфингомиелин, как правило, содержат только остатки насыщенных жирных кислот.
Мембранные фосфолипиды неравномерно распределены в наружном и внутреннем слоях (листках) бислоя. Существует своеобразная фосфолипидная асимметрия мембран. Она проявляется в том, что в наружном монослое мембраны в большем количестве, чем во внутреннем, содержатся фосфатидилхолин и сфингомиелин, а во внутреннем, в большем количестве чем в наружном, содержатся фосфатидилэтаноламин, фосфатидилсерин и фосфатидилинозитол. Подобная асимметрия имеет глубокий биологический смысл: более устойчивые к окислению фосфолипиды, содержащие насыщенные жирнокислотные остатки, находятся снаружи мембраны. Именно они в большей мере контактируют с воздействующими на нее агрессивными факторами внешней среды. Во внутреннем же листке бислоя, прямо не контактирующем с внеклеточной средой, находятся более чувствительные к окислению фосфолипиды, содержащие остатки ненасыщенных жирных кислот.
Помимо фосфолипидов, в состав липидного бислоя биологических мембран входят стерины. В животных клетках особое распространение среди них имеет холестерин. Молекула холестерина является полярной. На ней выделяется небольшой гидрофильный участок, представленный гилроксильной группой в третьем положении. Остальная часть молекулы – гидрофобна. Стериды, как правило, не входят в состав биологических мембран.
Содержание холестерина в мембранах невелико. Молярное соотношение фосфолипидов и холестерина (ФЛ/ХЛ) в них в среднем около 10 : 1. В то же время в мембранах эритроцитов отношение ФЛ/ХЛ достигает 1, а во внутренней мембране митохондрий холестерин вообще отсутствует.
Холестерин в липидном бислое образует многочисленные слабые связи с молекулами фосфолипидов и практические не взаимодействует с белками. Различные фосфолипиды существенно отличаются по сродству в отношении образования комплексов с холестерином. Наибольшее сродство к холестерину проявляют сфингомиелин и фосфатидилэтаноламин. Обнаружено, что существует определенное взаимоотношение между концентрацией холестерина и сфингомиелина в биологических мембранах.
Холестерин определенным образом ориентирован в плоскости мембраны. Он располагается перпендикулярно бислою (рис. 47). Его полярная гидроксильная группа находится в контакте с полярными головками молекул фосфолипидов. Остальная часть молекулы холестерина контактирует с жирнокислотными остатками фосфолипидов в центральной части бислоя. Взаимодействие между холестерином и фосфолипидами осуществляется преиму-щественно за счет Ван-дер-Ваальсовых сил. Во взаимодействии участвуют молекула холестерина и 1–2 молекулы фосфолипидов.

Рисунок 47 – Расположение холестерина в липидном бислое мембраны
(молекула холестерина выделена черным цветом)
Холестерин играет важную роль в мембране. Он во многом определяет физическое состояние липидного бислоя, выполняя функцию своеобразного конденсирующего агента, повышающего его плотность. Жирнокислотные остатки фосфолипидов, располагающиеся в центральной части липидного бислоя, подвержены определенной подвижности. Случайные движения одной из цепей передаются другим. Молекула же холестерина, встроенная между ними, блокирует подобное движение, предупреждая тем самым случайные нарушения упорядоченности жирнокислотных радикалов.
Помимо фосфолипидов и стеринов, в составе биологических мембран встречаются свободные жирные кислоты. Их концентрация в липидном бислое невелика. Однако при определенных состояниях уровень свободных жирных кислот в мембранах может существенно возрастать. Этот сдвиг приводит к изменению физических свойств мембраны и, как следствие того – к нарушению ее функционирования.
Липидный бислой мембраны представляет собой динамическую структуру, имеющую консистенцию оливкового масла. В нем происходит постоянное движение липидных молекул. Существуют две основные формы движения липидов в мембране:
1) латеральная диффузия, представляющая собой перемещение молекулы в плоскости бислоя;
2) движение типа flip-flop (перескок из одного монослоя в другой).
Преимущественной формой движения липидов в мембране является литеральная диффузия. Перескок липидной молекулы из одного листка бислоя в другой идет с незначительной скоростью. Это связано с тем, что подобная форма движения является энергозависимой.
Несмотря на то, что молекулы липидов находятся в мембране в сос-тоянии диффузии, липидный бислой не представляет собой гомогенного раствора липидов. По существующим представлениям он имеет жидкокристаллическую структуру, тоесть часть липидов в мембране находится в твердом агрегатном состоянии, а часть – в жидком. С помощью современных физических методов исследований показано, что в липидном бислое содержатся своеобразные образования – домены, имеющие кристаллическую структуру, которые “плавают” в жидких липидах. Домены состоят из насыщенных фосфолипидов (сфингомиелина, фосфатидилхолина) и стеринов, имеющих более высокую температуру плавления, чем основная масса мембранных фосфолипидов.
Соотношение жидкого и кристаллического компонентов липидного бислоя во многом зависит от жирнокислотного состава фосфолипидов. Это связано с тем, что включение в состав фосфолипидов остатков ненасыщенных жирных кислот приводит к понижению температуры их плавления. Повышение концентрации ненасыщенных жирнокислотных радикалов обусловливает понижение доли кристаллического компонента и, таким образом, повышение жидкостности (текучести) липидного бислоя мембраны. Изменение жидкокристаллической структуры мембраны оказывает выраженное влияние на состояние ее проницаемости. При понижении доли кристаллического компонента, т.е. при повышении ее жидкостности, происходит повышение мембранной проницаемости.
Следующим важным компонентом биологических мембран являются белки. Мембранные белки выполняют роль ферментов, рецепторов, ионных насосов и ионных каналов, преобразователей энергии и т.д. По этой причине именно они определяют специфические функции мембран.
Содержание белков в мембранах разного происхождения сильно различается. В состав большинства мембран входит около 40 % белков. Вместе с тем, величина этого показателя может существенно варьироваться. Качественный состав мембранных белков тоже отличается высокой вариабельностью. Так, в состав миелиновых мембран входят всего три разных белка, в мембранах эндоплазматического ретикулума мышечных клеток около 70 % всех белков представлено Са-зависимой АТФазой, а во внутренней митохондриальной мембране содержится около сотни разных белков. В большинстве случаев в состав мембран клеточных структур входят несколько десятков различных белков.
По характеру расположения в мембране мембранные белки подразделяются на две основные группы (рис. 48).


а б
Рисунок 48 – Расположение в мембране поверхностных (а)
и внутренних (б) белков
1. Внешние(периферические) белки. Эти белки легко освобождаются из мембраны при помощи экстракции буферными растворами определенной ионной силы. Как правило, они обратимо связаны с поверхностью мембраны с помощью сил слабых взаимодействий. По этой причине поверхностные белки свободно отделяются от мембраны;
2. Внутренние(интегральные) белки. Эти белки встроены внутрь липидного бислоя или насквозь проходят через толщу мембраны. Они жестко связаны с мембраной и могут быть выделены из нее только с помощью детергентов, разрушающих липидный бислой. Выделенные из мембраны внутренние белки, как правило, утрачивают характерные для них свойства и биологическую активность.
По аминокислотному составу мембранные белки не отличаются от немембранных (так называемых цитоплазматических). Однако при сравнении аминокислотного состава поверхностных и интегральных белков можно обнаружить, что интегральные белки содержат в своем составе гораздо больше аминокислот с гидрофобными радикалами (фенилаланин, аланин, лейцин, валин и др.). Подобное различие легко объясняется тем, что внутренние белки мембран находятся в прямом контакте с ее гидрофобным слоем.
Существуют особенности распределения аминокислот с неполярными и полярными радикалами в полипептидной цепи интегрального белка. Оказывается, что аминокислоты с неполярными радикалами сконцентрированы в той части полипептидной цепи, которая проникает в толщу липидного бислоя (рис. 48). За счет этого между неполярными аминокислотными радикалами и жирнокислотными остатками фосфолипидов возникают многочисленные гидрофобные связи, которые жестко стабилизируют белок внутри мембраны.
В верхней части рис. 49 обозначено положение полипептидных цепей в мембране. В нижней части рисунка изображена развернутая полипептидная цепь молекул белка. Положение аминокислот с неполярными радикалами вы-делено жирной линией.

Рисунок 49 – Особенности распределения аминокислот с неполярными
радикалами в полипептидной цепи интегральных (внутренних)
белков мембраны (а, б)
Подобно молекулам липидов мембранные белки находятся в состоянии постоянного движения в плоскости мембраны, взаимодействуя при этом друг с другом. Скорость движения белков в мембране во многом определяет интенсивность тех процессов, в которых данные белки принимают участие.
На подвижность белковых молекул в мембране большое влияние оказывает текучесть ее липидного бислоя. По этой причине липидный состав мембраны выступает в качестве одного из основных факторов, определяющих состояние мембранных процессов.
Некоторая часть мембранных белков обладает каталитическими свойствами, т.е. представляет собой ферменты. Выделяется особая группа так называемых мембраносвязанных ферментов. После удаления этих энзимов из мембраны они утрачивают свои каталитические свойства. При их повторном включении в структуру мембраны они реактивируются и вновь приобретают способность катализировать характерные для них реакции.
Реактивация мембраносвязанных ферментовможет быть достигнута и за счет добавления к ним суспензии некоторых фосфолипидов. При этом фосфолипиды выступают в роли активаторов, без которых проявление каталитических свойств ферментов невозможно.
Важное значение в регуляции активности мембранных ферментов имеет физическое состояние липидного бислоя и прежде всего его текучесть. При увеличении текучести липидов бислоя возрастает скорость диффузии белков в плоскости мембраны. При этом повышается эффективность их взаимодействия с субстратами, вследствие чего повышается и скорость катализируемых ими реакций.
Помимо липидов и белков, в состав большинства биологических мембран входят углеводы. Чаще всего углеводы встречаются в наружной клеточной мембране (плазмалемме).
Углеводы входят в состав мембран в связанном состоянии в форме гликолипидов или гликопротеидов. В их состав входят и остатки различных моносахаридов и их многочисленные производные: глюкоза, галактоза, манноза, глюкозамин, галактозамин, N-ацетилглюкозамин, N-ацетил-галактозамин, нейраминовая и сиаловые кислоты и др.
Гликопротеиды и гликолипиды содержат углеводный компонент, представленный ветвящимися олигосахаридными цепями, которые присоединяются к полипептидным цепям белком с помощью О- и N-глико-зидных связей или к липидам с помощью О-гликозидных связей.
Электронномикроскопические исследования показали, что углеводные компоненты располагаются на внешней поверхности наружной клеточной мембраны (рис. 50). Они могут быть легко удалены из нее путем обработки мембраны протеолитическими ферментами.

Рисунок 50 – Расположение гликопротеида в наружной
клеточной мембране
На поверхности многих типов животных клеток олигосахаридные цепи мембранных белков образуют сложную ветвистую структуру – гликокаликс. Гликокаликс играет важную роль в межклеточном взаимодействии, обеспечивает узнавание биологически активных веществ и их связывание, принимает участие в транспорте катионов через мембрану и многих других процессах.
При формировании представлений о целостной структуре биологических мембран создавались различные их модели. В настоящее время принята жидкостно-мозаичная модель G. Nickolson и С. Singer (1973 г.). Согласно этой модели биологические мембраны представляют собой трехмерные раст-воры белков в липидах (рис. 51).
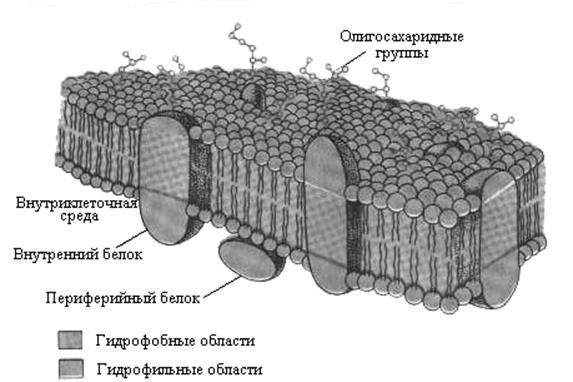
Рисунок 51 – Жидкостно-мозаичная модель мембраны
(А. Ленинджер, 1985)
В основе жидкостно-мозаичной модели мембран лежат следующие основные положения:
1) большая часть мембранных липидов принимает участие в образовании липидного бислоя, который выполняет двойственную роль – одновременно является растворителем для белков и формирует барьер проницаемости;
2) некоторая часть мембранных фосфолипидов связана с белками и необходима для их функционирования;
3) мембранные белки способны свободно диффундировать в плоскости липидного бислоя.
Несмотря на общность структуры, состав биологических мембран из разных источников может значительно различаться. Существенные различия характерны и для различных мембранных субклеточных структур внутри клетки.
Среди внутриклеточных мембран наименее всего изучены ядерные. Это связано со сложностью их выделения и очистки. Ядерные мембраны играют важную роль в клетке. Они регулируют ядерно-цитоплазматичес-кие взаимоотношения.
В отличие от других внутриклеточных мембран ядерная мембрана содержит большие отверстия – поры, хорошо проницаемые для макромолекул (белков и нуклеиновых кислот). Кроме того в ней отсутствует электрический потенциал.
Для ядерных мембран характерно примерно одинаковое содержание липидов (37–40 %) и белка (33–50 %). Фосфолипиды в них представлены в основном фосфатидилхолином и фосфатидилэтаноламином. Среди белков ядерной мембраны особое значение имеют фермент АТФаза, а также целый ряд окислительно-восстановительных энзимов.
Особые внутриклеточные структуры – митохондрии имеют два типа мембран – наружную и внутреннюю. Эти мембраны сильно различаются по составу, свойствам и функции. Наружная митохондриальная мембрана имеет ровные очертания. В отличие от нее внутренняя митохондриальная мембрана имеет многочисленные складки (кристы).
В состав липидного бислоя внутренней митохондриальной мембраны входит большое количество фосфолипида – кардиолипина, который в других мембранах вообще не встречается. В отличие от других внутриклеточных мембран эта мембрана не содержит сфингомиелина и холестерина. Содержание белков во внутренней митохондриальной мембране достигает 75 % от ее массы. При этом до 40–50 % белка внутренней митохондриальной мембраны приходятся на окислительно-восстановительные ферменты. Структурные белки мембран митохондрий обладают способностью к агрегации за счет резко выраженной гидрофобности.
Внутренняя митохондриальная мембрана хорошо проницаема для воды и небольших нейтральных молекул, но не проницаема для катионов Na+, K+, анионов Cl-, а также веществ, молекулярная масса которых превышает 50 Да. В этой связи во внутренней митохондриальной мембране содержится много белков-переносчиков, осуществляющих направленный транспорт веществ внутрь митохондрий и в обратном направлении.
Наружная митохондриальная мембрана богаче внутренней фосфолипидами. По фосфолипидному составу она сходна с мембраной эндоплазматического ретикулума. В ней имеются поры, через которые могут свободно проникать макромолекулы.
Причины различий в строении различных внутриклеточных мембран тесно связаны с особенностями их синтеза (биогенеза). Биогенез мембран представляет собой очень сложный и до конца не изученный процесс. Он происходит путем самосборки из их отдельных компонентов (липидов, белков и углеводов).
Биосинтез мембранных фосфолипидов и холестерина происходит на эндоплазматическом ретикулуме клетки. Синтезированные липидные молекулы связываются с соответствующими транспортными белками (фосфолипидобменивающими или холестеринтранспортными белками), переносятся через цитозоль к соответствующим мембранным структурам клетки (митохондриям, ядерным мембранам, лизосомам, пероксисомам, наружной клеточной мембране и др.) и встраиваются в них.
Синтез мембранных белков происходит на рибосомах в цитозоле клетки или на рибосомах, связанных с мембранами шероховатого эндоплазматического ретикулума. Синтезированные белки транспортируются к соответствующим внутриклеточным мембранам и встраиваются в них.
Аналогичным образом углеводные компоненты мембран образуются в эндоплазматическом ретикулуме и комплексе Гольджи, после чего связываются с белками или липидами и уже в таком виде используются для построения мембран.
Контрольные вопросы
1. Почему клеточная мембрана в поперечном разрезе на электронных микрофотографиях имеет характерную трехслойную структуру?
2. Какие методы фракционирования клеточных мембран вы знаете? Что положено в их основу?
3. Чем представлен липидный бислой мембран?
4. Почему именно фосфолипиды представляют собой основной структурный компонент липидного бислоя мембраны?
5. Какие фосфолипиды доминируют в биологических мембранах?
6. В комплексе с какими фосфолипидами в мембранах присутствует холестерин?
7. Чем обусловлена жидкокристалическая структура липидного бислоя мембраны?
8. В чем заключается фосфолипидная ассиметрия липидного бислоя мембраны?
9. Какие факторы определяют проницаемость липидного бислоя мембраны?
10. Что вы можете сказать о структурных особенностях мембранных и не мембранных белков?
11. Что влияет на скорость диффузии мембранных белков?
12. Какие характерные особенности присущи мембраносвязанным белкам? Приведите примеры мембраносвязанных белков.
13. Каким образом располагаются в мембране углеводные компоненты?
14. Как происходит внутриклеточное образование мембран?
Глава 10. ГОРМОНЫ
Живые существа обитают в условиях постоянно меняющейся внешней и внутренней среды. При этом возникает реальная необходимость в своевременной коррекции у них состояния обменных процессов, что позволяет организму приспосабливаться к новым условиям существования. Важную роль в подобном приспособлении играют особые биологически активные вещества – гормоны.
В классическом представлении – гормоны это биологически активные вещества, образующиеся в эндокринных железах, секретируемые в кровь и оказывающие регуляторное влияние на обмен веществ и функцию внутренних органов. В настоящее время известно большое число гормонов животных и растений (фитогормонов).
Существуют различные классификации гормонов животных и человека. В их основу положены место образования и секреции, характер физиологического эффекта и др. По месту секреции гормоны подразделяются на:
1) гормоны гипоталамуса (рилизинг-факторы, статины);
2) гормоны передней доли гипофиза (адренокортикотропный гормон (АКТГ), соматотропный гормон, фолликулостимулирующий гормон, лютеинизирующий гормон, лютеотропный гормон, тиреотропный гормон);
3) гормоны задней доли гипофиза (вазопрессин, окситоцин);
4) гормоны щитовидной железы (тироксин, трийодтиронин, кальцитонин);
5) гормоны паращитовидных желез (паратгормон);
6) гормоны поджелудочной железы (инсулин, глюкогон);
7) гормоны коркового (глюкокортикоиды, минералкортикоиды) и мозгового слоя (адреналин) надпочечников;
8) гормоны половых желез (тестостерон, эстрадиол);
9) гормоны шишковидной железы (мелатонин) и др.
Особое значение имеет классификация, основанная на различиях в химической структуре гормонов. По этой классификации все гормоны подразделяются на следующие группы:
1) пептидные гормоны (к ним относятся гормоны пептиды, а также простые и сложные белки);
2) гормоны – производные аминокислот (адреналин, тиреоидные гормоны и др.);
3) стероидные гормоны (производные холестерина).
Гормоны пептидной природы могут образовываться в неактивном виде – в форме прогормонов (инсулин, глюкогон и др.). В таком неактивном виде они, как правило, накапливаются в ткани железы. Однако секретируется гормон уже в активном виде. Процесс образования активного гормона обычно связан с его частичным протеолизом, в результате которого происходит уменьшение его молекулярной массы.
Уровень гормонов в крови в покое невелик и соответствует микромолярной (10-6 М), наномолярной (10-9 М) или пикомолярной (10-12 М) концентрации. В условиях стимуляции функции эндокринной функции железы уровень гормонов может возрастать в 100 и более раз.
Время жизни большинства известных гормонов невелико и составляет секунды. Это связано с тем, что они очень быстро распадаются под влиянием различных ферментов крови. Небольшая продолжительность существования, а значит, и проявления их регуляторного эффекта, позволяет гормонам участвовать в формировании срочных регуляторных влияний на организм, действие которых соответствует продолжительности влияния неблагоприятного фактора. В то же время некоторые гормоны (стероидные, тиреоидные) оказывают продолжительное влияние на организм. За счет этого они обеспечивают возникновение долговременной адаптации к действию неблагоприятных факторов.
Гормоны оказывают влияние не на все, а лишь на определенные органы, которые получили название “органы-мишени”. Для каждого гормона существуют свои органы-мишени. Их количество для каждого гормона существенно различается. Одни из них имеют только один орган-мишень (тиреотропный гормон – щитовидная железа, АКТГ – кора надпочечников и др.), другие – несколько (органами-мишенями инсулина являются печень, скелетные мышцы, жировая ткань и др.)
Существование органов-мишеней обусловлено присутствием в них особых, воспринимающих гормоны, белковых структур – рецепторов. Рецепторы представляют собой отдельные белки или белковые комплексы, способные специфически связывать гормоны, в результате чего происходит возникновение ответа со стороны клеток органов-мишеней.
Рецептор состоит из двух различных доменов (участков):
1) участка узнавания и связывания гормона;
2) участка генерации специфического внутриклеточного сигнала, обеспечивающего взаимосвязь процесса присоединения гормона к рецептору с клеточным ответом.
Связывание гормона рецептором происходит за счет структурного соответствия гормона или его определенного участка и участка связывания рецептора. В результате связывания происходит образование гормон-рецепторного комплекса, который стабилизируется многочисленными силами слабых взаимодействий – водородными связями, силами электростали-ческого и гидрофобного взаимодействия, Ван-дер-Ваальсовыми силами. Возникающий комплекс непрочен и способен самопроизвольно распадаться.
Количество рецепторов и их сродство к гормону является непостоянными величинами. В разных условиях они могут существенно изменяться.
Гормональные рецепторы могут располагаться в различных отделах клетки органа-мишени. Для пептидных гормонов и гормонов производных аминокислот они располагаются на внешней поверхности клеточной мембраны. Рецепторы же стероидных и тиреоидных гормонов локализуются в цитоплазме клетки.
В зависимости от внутриклеточной локализации рецепторов все гормоны подразделяются на:
1) гормоны, взаимодействующие с внутриклеточными рецепторами;
2) гормоны, взаимодействующие с мембранными рецепторами.
К гормонам первой группы относятся стероидные (гормоны половых желез и коры надпочечников) и тиреоидные (тироксин, трийодтиронин) гормоны, а ко второй – адреналин и все пептидные гормоны. Гормоны этих групп обладают принципиально различным механизмом регуляторного эффекта на клетки органов-мишеней.
|
Просмотров 4873 |
|
|
